

Sturia brandti n. sp. und Sephardonautilus nov. gen.
Die Arbeit untersucht den Oberen Muschelkalk als neu entstandenes Binnenmeer-Ökosystem der Mittleren Trias und zeigt, wie Einwanderungen von Ammonoideen aus alpinen und sephardischen Faunenprovinzen biostratigraphische und chronostratigraphische Korrelationen ermöglichen. Anhand neu beschriebener Taxa (Sturia brandti n. sp. - entdeckt von Sebastian Brandt und Sephardonautilus nov. gen.) werden Migrationswege, allopatrische Speziationsprozesse und evolutionäre Zusammenhänge rekonstruiert. Die Ergebnisse liefern eine präzisere zeitliche Gliederung des Oberen Muschelkalks und neue Einblicke in die Faunendynamik während des Anisium/Ladinium-Übergangs.
Sigfried Rein, Sebastian Brandt


Sturia brandti n. sp. und Sephardonautilus nov. gen. – Immigranten, Migrationswege und Korrelationen im Oberen Muschelkalk (Mittlere Trias)
Zusammenfassung: Als neu entstandenes Ökosystem ist der Obere Muschelkalk ein tektonisch dominiertes Binnenmeer. Immigrierte Makrofossilien aus den alpinen und sephardischen Faunenprovinzen ermöglichen die biostratigraphische und chronostratigraphische Korrelation. In der unteren trinodosus-Zone migrierte Germanonautilus salinarius aus der austroalpin-dinarischen Faunenprovinz über eine helvetisch-alemannische Pforte in das neu entstandene Binnenmeer. Aus der Stammart entstand kladogenetisch Germanonautilus bidorsatus. Auf dem gleichen Weg migrierte in der mittleren trinodosus-Zone Sturia sansovinii aus der austroalpinen Faunenprovinz und bildete kladogenetisch die neue Spezies Sturia brandti n. sp.
Der bedeutendste Schritt der Besiedlung des Ökosystems Oberer Muschelkalk erfolgte im unteren Illyrian aus der sephardischen Faunenprovinz durch eine einmalig kurzzeitig tektonisch geöffnete burgundisch-rheinische Pforte. Mit Sephardonautilus tridorsatus nov. comb. als Typusart der neuen Gattung Sephardonautilus nov. gen. und der Stammart Sephardonautilus salinarius wird die verwandtschaftliche Verbundenheit zwischen den Individuen des Muschelkalkmeeres und der sephardischen Faunenprovinz dokumentiert. Gleiches gilt für die nach allopatrischer Speziation aus einer unbekannten Art der Gattung Ceratites de Haan der sephardischen Faunenprovinz entstandene evolutionäre Art Ceratites nodosus.
Die Stammart Sephardonautilus salinarius aus der trinodosus-Zone der oberen Gevanim-Formation von Makhtesh Ramon belegt chronologisch die Korrelation des Migrationsweges des Sephardonautilus tridorsatus. Die Morphokline der beispiellos eurypotenten evolutionären Art Ceratites nodosus ermöglicht die lückenlose biostratigraphische Gliederung des Oberen Muschelkalkes.
Populationen der südalpinen Morphen Flexoptychites angusto-umbilicatus und Protrachyceras recubariense immigrierten hypothetisch zeitgleich mit dem Brachiopoden Punctospirella fragilis. Der Zeitpunkt dieser vierten Immigrationsphase entsprach chronostratigraphisch und biostratigraphisch dem Anisian/Ladinian-Wechsel. Mit physiologischer Anpassung überleben Flexoptychites und Protrachyceras in SW-Deutschland von der evolutus-Zone bis in die spinosus-Zone. Sie erwarten einen neuen kladogenetischen Artstatus.
Für die Festlegung des Anisian/Ladinian-Wechsels im Oberen Muschelkalk ergäbe sich die Spiriferina-Bank mit dem Wechsel von der compressus-Zone zur evolutus-Zone. Er korreliert in der südalpinen Faunenprovinz mit dem Wechsel von der secadensis-Zone zur curionii-Zone.
Die Immigranten aus dem Ladin der Schreyeralm-Formation Parapinacoceras thiemei und Gymnites bruneli besiedeln kurzzeitig das Beckeninnere in Thüringen. Durch die allopatrische Speziation einer unbekannten Spezies der Gattung Ceratites de Haan aus der sephardischen Faunenprovinz entstanden die Schwesterarten Ceratites nodosus und Ceratites tornquisti. Die Phylogenese der Spezies Ceratites tornquisti beginnt im unteren Illyrian in der Paläotethys und endet im Longobardian im Thüringer Grenzdolomit als Chronospezies Ceratites schmidti. Die Klärung der bislang unbekannten Phylogenese von Ceratites tornquisti in den Lebensräumen Dobruscha, Sardinien, Vicentin, Provence und Thüringen bleibt einer detaillierten Analyse vorbehalten.
1. Einleitung
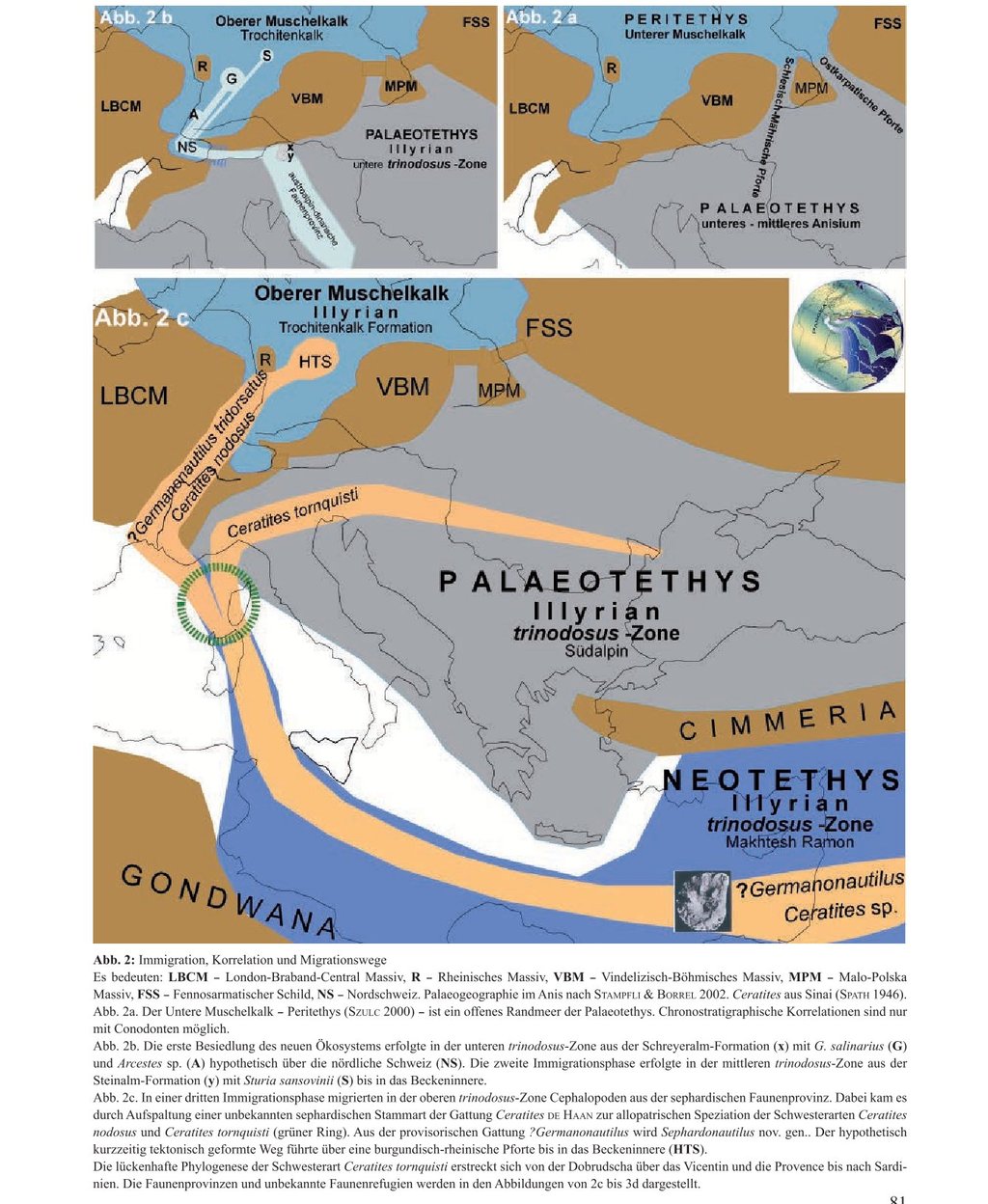
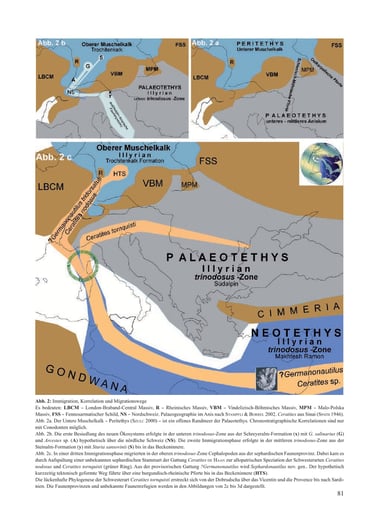
In der Mitteltrias war das heutige Mitteleuropa ein im Südosten durch das Böhmisch-Vindelizische Hochland und das Karpatenvorland vom Paläotethys-Ozean getrennter Senkungsraum. Epirogenetisch und eustatisch verursacht wurde er durch die Ostkarpatische Pforte und später durch die Schlesisch-Mährische Pforte geflutet (Abb. 2a). So entstand im Unteren Muschelkalk als flaches Randmeer der Paläotethys die Peritethys (Szulc 2000). Mit einem nördlich geschlossenen Küstenrand erstreckte sie sich von Polen über Deutschland bis nach Frankreich und an den Nordrand der Schweiz. Bis zum mittleren Anisian erfolgte über diese Pforten die Faunenmigration ungehindert aus den asiatischen und alpinen Faunenprovinzen der Paläotethys (Kozur 1974). Die paläogeographische Position dieser Wanderwege aus der am Südostrand direkt anliegenden Paläotethys in dieses Randmeer ist relativ gut bestimmt.
Die fossilarmen Karbonate des Unteren Muschelkalks ermöglichen jedoch keine beckenweite Biostratigraphie mit Makrofossilien. Deshalb sind aufgrund fehlender biostratigraphischer Referenzprofile chronostratigraphische Korrelationen mit dem Tethys-Raum lediglich mithilfe von Conodonten möglich (Narkiewicz & Szulc 2004).
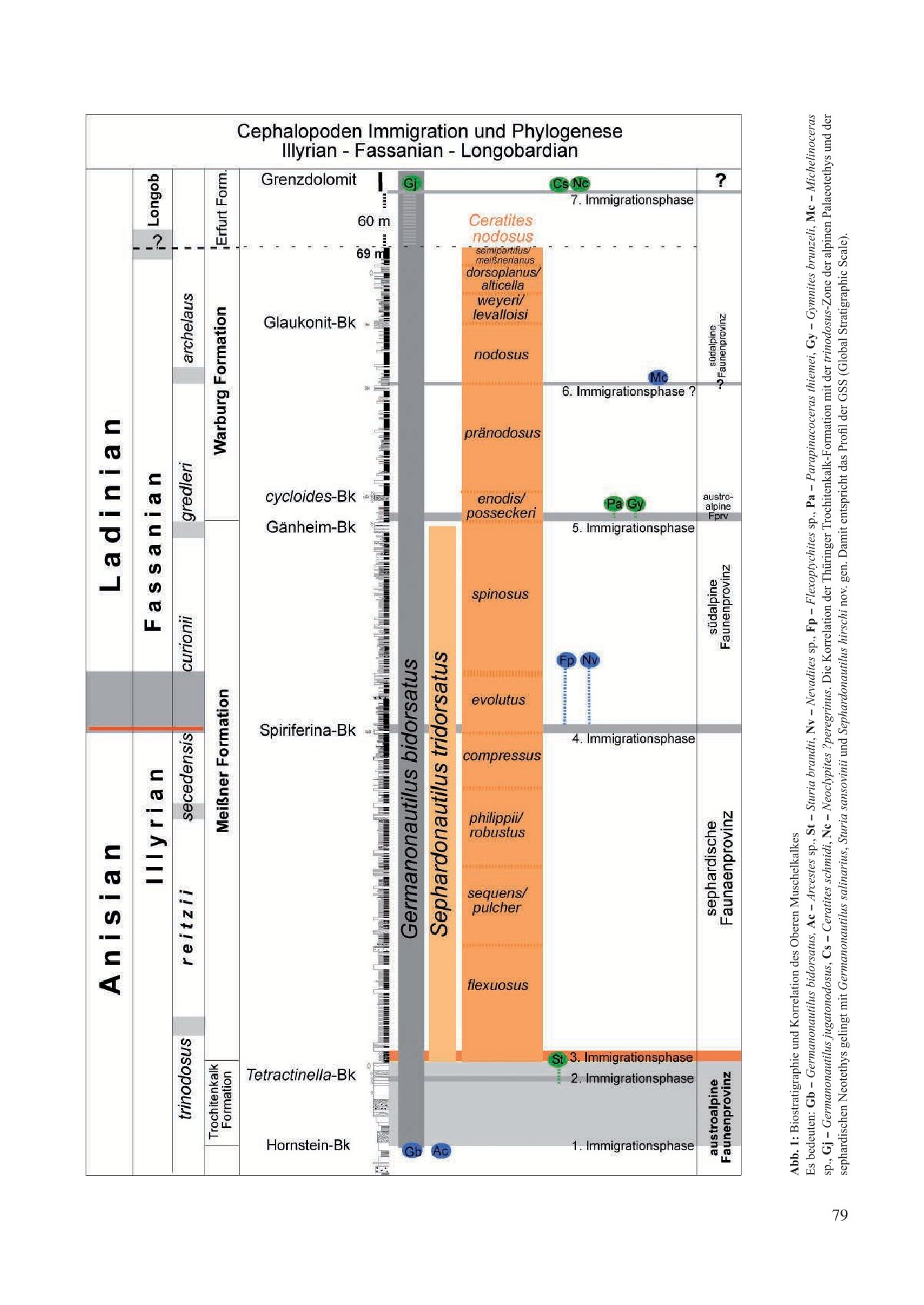
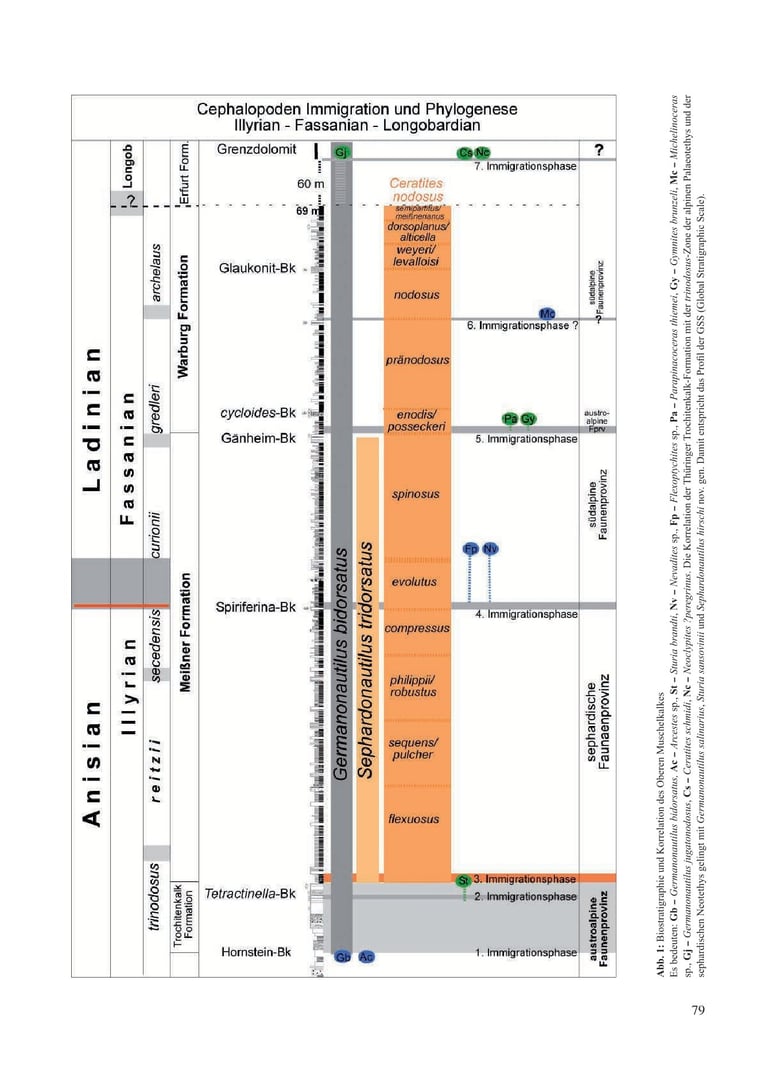
Nach der Schließung der östlichen Pforten wurde die marine Verbindung zur Paläotethys völlig unterbrochen. Mit dem durch erhöhte Verdunstung verursachten extremen Anstieg der prozentual im Meerwasser gelösten Salze erloschen im Mittleren Muschelkalk über 1,1 Ma Jahre weitgehend die Lebensbedingungen.
2. Der Obere Muschelkalk
Im Gegensatz zu den im Unteren Muschelkalk paläogeographisch relativ gut definierten östlichen Pforten eines Randmeeres bleiben im Oberen Muschelkalk alle Deutungen für Pfortenöffnungen zum Tethys-Ozean hypothetisch. Im Unterschied zum Unteren Muschelkalk sind die Karbonate des Oberen Muschelkalkes jedoch ungewöhnlich fossilreich. Sie garantieren mit Makrofossilien die Erstellung einer hochauflösenden Biostratigraphie und ermöglichen eine chronologische Dokumentation des neu entstandenen Ökosystems der Germanischen Faunenprovinz. Die evolutionäre Entwicklung der Organismen erfolgte nach ihrer Immigration unter endemischen Bedingungen. Die Immigration einer Art endete mit reproduktiver Isolation von ihrer Stammart und der kladogenetischen Entstehung einer neuen Spezies.
Das biostratigraphische Profil belegt mit Makrofossilien sieben Immigrationsphasen in dieses Binnenmeer und damit neue Artbildungen (Abb. 1). Als chronologische Dokumentation ermöglichen sie die Korrelation mit verschiedenen tethyalen Faunenprovinzen und die Rekonstruktion hypothetischer Wanderwege der Immigranten.
Als Schinznach-Formation ist in der Nordschweiz der Obere Muschelkalk vollständig ausgebildet (Pietsch et al. 2016; Adams et al. 2019). Makrofossilien belegen die südwestlichste Ausdehnung des Muschelkalkmeeres (Merki 1961). Ganz im Osten der Schweiz liegt das St. Galler Rheintal. Es grenzt an Österreich und Liechtenstein. Eine Verbindung über die Alpenrhein-Depression zum Westrand der Paläotethys erscheint deshalb geologisch denkbar („Western Gate“, Szulc 2000). Sie würde den kürzesten Wanderweg am östlichen Schwarzwald-Rand durch Südwestdeutschland in das Becken bedeuten („Alemannische Pforte“, Bachmann et al. 2010).
Favorisiert wurde bislang eine marine Verbindung nach Süden über eine „Burgundische Pforte“. Zumindest die aus der sephardischen Faunenprovinz der Neotethys in das Muschelkalkmeer immigrierten Cephalopoden müssten diesen burgundischen Weg zwischen den Vogesen und dem westlichen Schwarzwaldrand durch eine „Rheinische Pforte“ benutzt haben (Rein 2016).
3. Die Immigrationsphasen
3.1 Erste Immigrationsphase (Abb. 1, Abb. 2b/x)
Mittlerer Muschelkalk, Diemel-Formation
Rhyncholithes hirundo (Germanonautilus) (Gb): Fundort Oßmaritz bei Jena, Thüringen, Liegendes der Oberen Hornsteinbank (Knaust 1991).
Arcestes sp. (Ac): Fundort Weil der Stadt vom östlichen Schwarzwaldrand, Südwestdeutschland, Liegendes der Hornsteinbank (Hohenstein 1913; Schmidt 1928).
3.2 Zweite Immigrationsphase (Abb. 1, Abb. 2b/y)
Oberer Muschelkalk, Trochitenkalk-Formation, Tetractinella-Bank, Thüringen
Tetractinella trigonella: Ökostratigraphische Leitbank
Sturia sansovinii (St): Fundort Geilsdorf bei Stadtilm, Hangendes der Tetractinella-Bank, Thüringen.
3.3 Dritte Immigrationsphase (Abb. 1)
Oberer Muschelkalk, Meißner-Formation, flexuosus-Zone, Thüringen
Ceratites nodosus: Hessisch-Thüringische Senke (Seidel 2013), Thüringen, Troistedt, ca. 1 m über der Tetractinella-Bank (Rein 2007, 2017).
?Germanonautilus tridorsatus: Hessisch-Thüringische Senke, Thüringen, Troistedt, ca. 1 m über der Tetractinella-Bank (Rein 2016).
3.4 Vierte Immigrationsphase (Abb. 1)
Oberer Muschelkalk, Meißner-Formation, Spiriferina-Bank, Südwestdeutschland
Punctospirella fragilis: Ökostratigraphische Leitbank
Flexoptychites angusto-umbilicatus (N): Fundort Gundelsheim, spinosus-Zone (Urlichs & Kurzweil 1997).
Protrachyceras recubariense (P): Fundort Steinbruch am Winterrain bei Weissach, Südwestdeutschland, mittlere spinosus-Zone (Urlichs 1978).
3.5 Fünfte Immigrationsphase (Abb. 1)
Oberer Muschelkalk, Warburg-Formation, Thüringen
Pinacoceras sp. (Pa): Fundort Ettenhausen bei Eisenach, Thüringen, Warburg-Formation, enodis/posseckeri-Zone (Rein 2009; Rein & Werneburg 2010).
Gymnites sp. (Gy): Fundort Ohrdruf bei Gotha, Thüringen, Warburg-Formation, enodis/posseckeri-Zone (Rein & Werneburg 2010).
3.6 Sechste Immigrationsphase (Abb. 1)
Oberer Muschelkalk, Warburg-Formation, SW-Deutschland
Michelinoceras campanile (Mc): Fundort Trigonodus-Dolomit von Buigen bei Mönsheim, SW-Deutschland, stammt aufgrund der Begleitfauna vermutlich aus der nodosus-Zone (Schröder 1977; Urlichs & Schröder 1980).
3.7 Siebente Immigrationsphase (Abb. 1)
Unterer Keuper, Erfurt-Formation, Grenzdolomit, Thüringen
Ceratites schmidti (Cs): Fundorte Sülzenbrücken und Reisdorf, Thüringen, Grenzdolomit (Zimmermann 1883; Philippi 1901; Müller 1969, 1970, 1973).
Neoclypites? peregrinus (Nc): Fundort Reisdorf, Thüringen, Grenzdolomit (Müller 1973).Germanonautilus jugatonodosus (Jn): Branchewinda, Thüringen, Grenzdolomit (Zimmermann 1889; Rein 2014, 2015; Urlichs & Mundlos 1984; Urlichs 2000, 2015).
Beschreibung
4.1 Immigration von Germanonautilus salinarius aus der austroalpin-dinarischen Faunenprovinz (Schreyeralp-Formation) (Abb. 2b/x)
In der Mitteltrias erstreckt sich mit gemeinsamer Fauna von Han Bulog bis zu den Schreyeralmkalken des Salzburger Landes die austroalpin-dinarische Faunenprovinz. Die Schreyeralp ist die Typlokalität von Nautilus salinarius Mojsisovics, 1882 und Arcestes sp. Sie belegt mit Paraceratites trinodosus ein illyrisches Alter. Nautilus salinarius wird auch von der Schiechlinghöhe (Diener 1900) und Han Bulog (Hauer 1887) beschrieben.
An der Basis des Illyrian erfolgte in der unteren trinodosus-Zone auch die erste Immigrationsphase in den im Oberen Muschelkalk neu entstehenden Lebensraum. Die immigrierten Nautilus salinarius und Arcestes sp. aus der austroalpin-dinarischen Faunenprovinz ermöglichen die erste chronostratigraphische und biostratigraphische Korrelation mit der Paläotethys. Deskriptiv wurde aus Nautilus salinarius Mojsisovics, 1882 als Stammart die neue Spezies Nautilus bidorsatus Schlotheim, 1820. Beide Arten wurden von Mojsisovics (1902) in der neu gefassten Gattung Germanonautilus vereint und Germanonautilus bidorsatus als Typusart der neuen Gattung bestimmt.
Paläogeographisch ist der gewählte Wanderweg über die Alpenrhein-Depression hypothetisch. Mit der Ausbildung des Oberen Muschelkalks belegt die Schinznach-Formation in der Nordschweiz (Pietsch et al. 2016; Adams et al. 2019) jedoch die Möglichkeit einer marinen Verbindung über die Alemannische Pforte (Bachmann et al. 2010) nach SW-Deutschland.
Mit dem Fund des Rhyncholithes hirundo aus dem Mittleren Muschelkalk von Jena (Knaust 1991) wird die Immigration des Germanonautilus salinarius aus dem austroalpinen Raum der Paläotethys in der trinodosus-Zone belegt. Da der Kiefer ein Teil des Weichkörpers ist, kann ein postmortaler Transport ausgeschlossen werden. Somit ist sein Fund im Beckeninneren ein Beweis für die erfolgreiche Besiedlung des neuen Lebensraumes in der oberen Diemel-Formation. Er ist weiterhin ein physiologischer Beleg für die ungewöhnliche eurypotente Anpassungsfähigkeit des Nautiliden. Die nach seiner reproduktiven Isolation neu entstandene Spezies Germanonautilus bidorsatus bleibt die einzige alpine Cephalopoden-Spezies, die sich dauerhaft im Oberen Muschelkalk integrieren konnte.
Stratigraphisch beginnt der Obere Muschelkalk in SW-Deutschland und Thüringen mit der Hornsteinbank. Bislang wurde der Fund von Arcestes sp. aus dem Mittleren Muschelkalk SW-Deutschlands als postmortal passiv transportierte Schale gedeutet. Am Beispiel von Germanonautilus wird jedoch deutlich, dass die aktive Besiedlung des neuen Lebensraumes bereits in der Diemel-Formation mit dem ersten Frischwasser der transgredierenden Paläotethys einsetzte. Korreliert mit der Immigration des Germanonautilus salinarius entspricht dies der unteren trinodosus-Zone.
4.2 Immigration von Tetractinella trigonella und Sturia sansovinii aus der austroalpinen Faunenprovinz (Steinalm-Formation) (Abb. 2b/y)
Die Konstellation der Immigration des Brachiopoden Tetractinella trigonella und des Cephalopoden Sturia sansovinii unterscheidet sich von der Immigration des Germanonautilus salinarius. Tetractinella trigonella ist aus der Fazies der Schreyeralm-Formation (Schreyeralp, Schiechlinghöhe) nicht nachgewiesen (Bittner 1890). Bekannt ist Tetractinella trigonella aus der binodosus-Zone der östlichen Kalkalpen, dem Balaton-Hochland und Recoaro (Bittner 1890; Torti & Angiolini 1997; Palfy 2003). Bedeutsam erscheint daher der Nachweis von Tetractinella trigonella aus der trinodosus-Zone der Steinalm-Formation (Broili 1927).
Dagegen ist Sturia sansovinii ein durch seine weite horizontale Verbreitung wichtiges Leitfossil der trinodosus-Zone (Diener 1900). Belegt ist es aus der Typlokalität Schreyeralmkalk (Mojsisovics 1882), der Schiechlinghöhe (Diener 1900) sowie der Steinalm-Formation (Broili 1927; Schnetzer 1934).
Ein weiteres Beispiel für die weite horizontale Verbreitung als Leitfossil der trinodosus-Zone ist der Nachweis von Sturia sansovinii im Thüringer Muschelkalk. Der Fund wurde am 24. Juni 2016 beim Bau der Bundesstraße B 90 bei Geilsdorf (R 44 35 590; H 56 22 800) im Aushub gemacht. Mit diesem Fund wird die Korrelation der oberen Trochitenkalk-Formation in Thüringen mit der alpinen mittleren trinodosus-Zone bestätigt. Er besitzt daher einen besonderen Wert für die chronostratigraphische Korrelation zwischen der germanischen und alpinen mittleren Trias.
Der Fundort Geilsdorf liegt im Beckenzentrum; die Fundlage befindet sich stratigraphisch etwa 3,5 m über dem Immigrationsniveau von Tetractinella trigonella. Sturia sansovinii war mit hoher Wahrscheinlichkeit kein Einzelindividuum, sondern Teil einer Einwandererpopulation. Bedingt durch territoriale und reproduktive Isolation von der alpinen Stammart entstand unter endemischen Bedingungen eine neue kladogenetische Spezies.
Systematik
Familie Ptychitidae Mojsisovics, 1882
Gattung Sturia Mojsisovics, 1882
Typusart Sturia sansovinii Mojsisovics, 1882
Art Sturia brandti n. sp.
Beschreibung von Sturia brandti n. sp.
Holotypus: Einzelfund vom 24.06.2016, Fundort Geilsdorf (R 44 35 590; H 56 22 800), Oberer Muschelkalk, Trochitenkalk-Formation, Thüringen, Sammlung des Naturkundemuseums Erfurt, Sammlungs-Nr. NME C16;200.
Erhaltung: Es liegt ein halbseitig erhaltener Steinkernrest mit ca. 120° des Phragmokon-Umganges vor. Außer limonitischen Belägen auf der Steinkernoberfläche als diagenetisch umgewandelte Reste des Hypostracums sind keine Schalenreste fossil erhalten. Daher fehlen Oberflächenstrukturen der Lateralseite.
Derivatio nominis: Der Artname Sturia brandti n. sp. bezieht sich nicht nur auf den Finder des Fossils, sondern würdigt die vielseitigen Leistungen des im Thüringer Muschelkalk verdienstvoll tätigen Sebastian Brandt aus Kornhochheim.
Vergleich mit dem Original von Sturia sansovinii Mojsisovics, 1882
Größe: Die Breite des Fundes beträgt marginal 124 mm. Die Höhe der Windung nimmt von ca. 65 mm auf 86 mm zu.
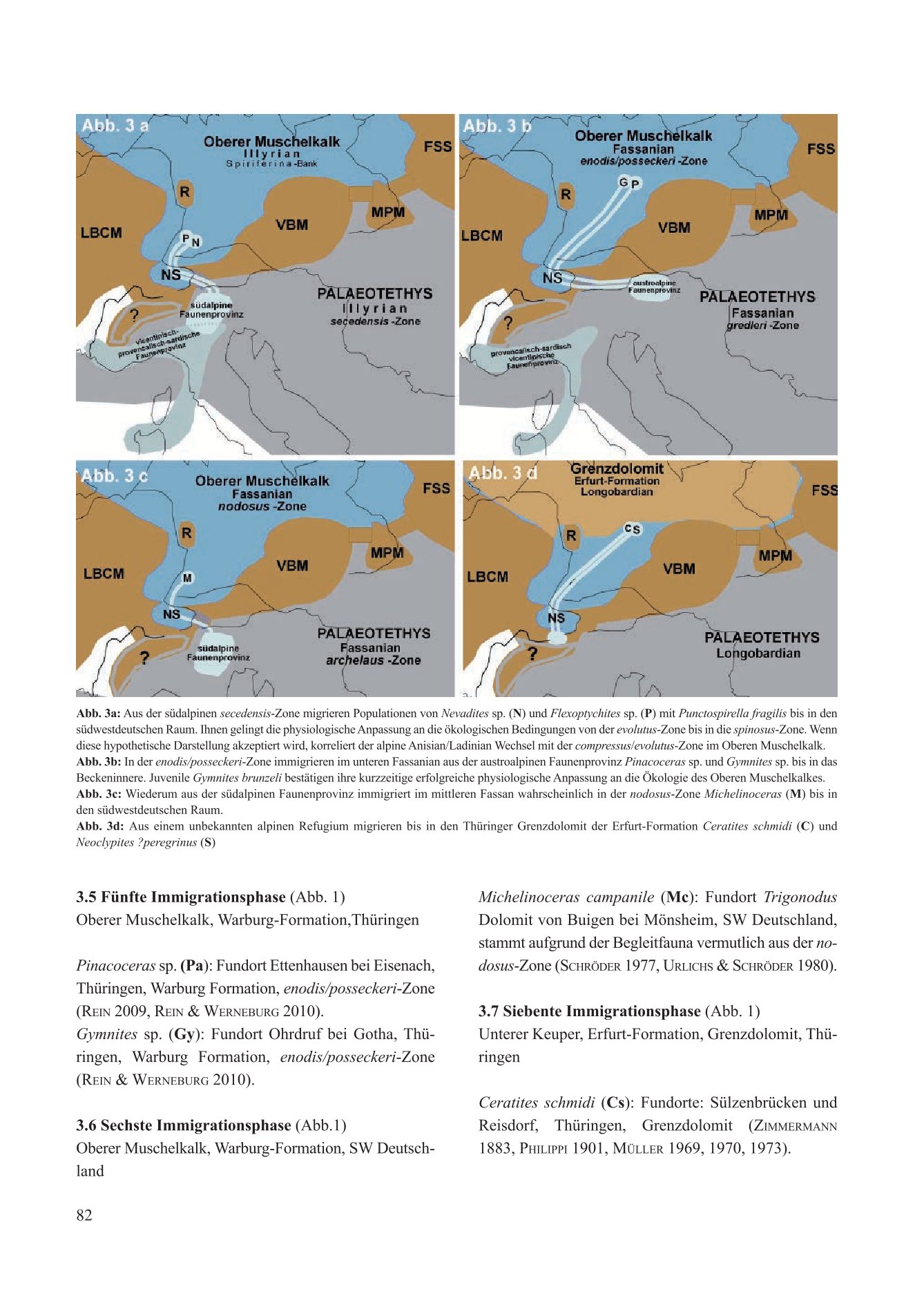
Abb. 4: Sturia brandti n. sp., Geilsdorf, untere Meißner-Formation Thüringen


Der Gehäusequerschnitt Q = (b/h)*100 kann nur angenähert berechnet werden. Er beträgt gleichbleibend 46. Die Werte der Windungshöhe des Originals betragen 68 mm und der Gehäusequerschnitt 41.
Mojsisovics (1882) beschreibt den Phragmokon-Durchmesser des größten ihm vorliegenden Exemplares mit 135 mm. Mit einem rekonstruierten Gesamt-Durchmesser von ca. 225 mm (Abb. 6) liegt die Größe des Thüringer Fundes geringfügig über den Funden der Schreyeralp-Morphen Mojsisovics.
Suturvergleich (Abb. 5) Abb. 5
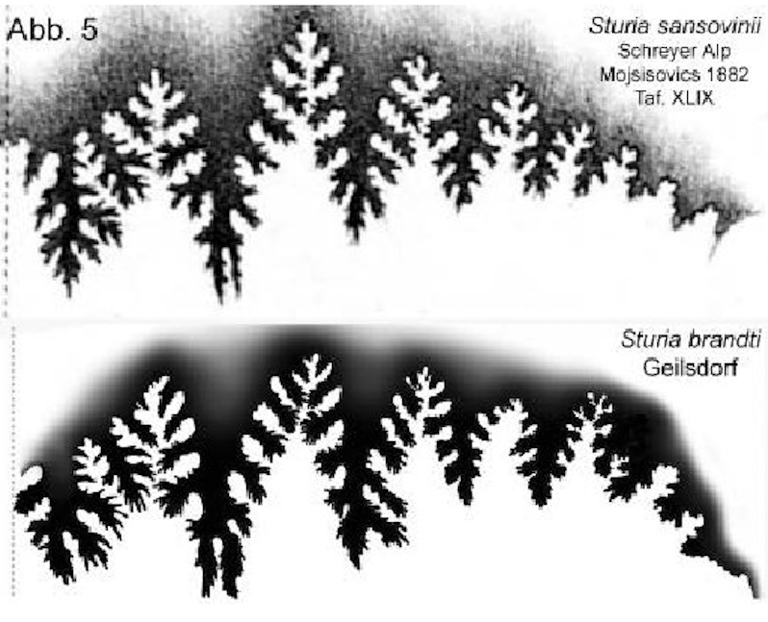


Abb. 6: Rekonstruktion der Gehäusegröße von Sturia brandti n. sp.
Abb. 5: Sutur-Vergleich von Sturia sansovinu Mojsisovics und Sturia brandti n. sp.
Loben: Es sind nur fünf Auxiliarloben ausgebildet. Der Externlobus wird durch einen breiten, auf die Seiten-flanken übergreifenden Medianhöcker geteilt. Der Ex-ternlobus ist breiter wie beim Original. Auch der erste und zweite Laterallobus spaltet zweispitzig deutlich breiter wie beim Original auf.
Sättel: Der Externsattel erscheint wie beim Original breit und besitzt einen deutlichen Außenast. Die Sättel sind schmal, hoch, zugespitzt, in der oberen Hälfte bis auf den schmalen Sattelstamm eingeschnitten und unten mit einem breiten, nicht zerschnittenen Sattelstamm versehen. Der Externsattel ist wie beim Original etwas niedriger als der erste Lateralsattel.
Die Sutur des Thüringer Fundes unterscheidet sich beim Vergleich mit dem Original von Sturia sansovinii Mojsisovics, 1882 deutlich bei der Ausbildung der Lateralloben und der Sattelstämme.
4.3. Die Immigration der sephardischen Cephalo-poden (Abb. 2c)
Die Cephalopoden der dritten Immigrationsphase im basalen Illyrian, ?Germanonautilus tridorsatus Bött-cher, 1938 und Ceratites nodosus Schlotheim, 1820, waren keine Arten aus dem alpinen Tethysraum sondern fremde Formen der Neotethys. Den realen Zeitpunkt der Immigration in das Muschelkalkmeer belegt der Fossilbericht (REIN 2008, 2016, 2017). Im Oberen Muschelkalk Thüringens erstreckt sich die Trochitenkalk-Formation (basales Illyrian) von der oberen Hornsteinlage bis einen Meter über die Tetractinella-Bank. Mit dem Fazieswechsel über dieser ökostratigraphischen Leitbank (HAGDORN & SIMON 1993) beginnt in Thüringen die Meißner-Formation und biostratigraphisch mit den ersten Ceratites die flexuosus-Zone. In den Ton/Mergel Lagen dieser Bio-zone erfolgt der erste gemeinsame fossile Nachweis der Cephalopoden ?Germanonautilus tridorsatus und Ceratites nodosus.
Er belegt ihre gemeinsame Immigration über die Rheinische Pforte (rein 2017) in die Hessisch-Thüringische Senke (Seidel 2013) und damit direkt in das Zentrum des Muschelkalkmeeres. Der Migrationsweg in die germanische Faunenprovinz ist hypothetisch (Abb. 2c). Er kann jedoch biostratigraphisch und chronostratigraphisch mit der genealogischen Korrelation des ?Germanonautilus tridorsatus nachvollzogen werden (rein 2016). Eine statistisch-biometrische Analyse ergab, dass die ursprünglich zur Gattung Germanonautilus Mojsisovics, 1902 gestellte Form Germanonautilus tridorsatus Böttcher, 1938 morphologisch und physiologisch zu einer eigenen Gattung gestellt werden müsste (rein 2016). Da der Ursprung der Stammreihe der Spezies ?Germanonautilus tridorsatus Böttcher, 1938 in der sephardischen Faunenprovinz liegt, wurde vorerst der Gattungsbegriff ?Germanonautilus gewählt.
Von Kummel (1960) wurden die nautiliden Morphen aus Areif en Naqa und Makhtesh Ramon (Sinai/Negev) mit steilem Nabeleinfall zur Gattung Germanonautilus Mojsisovics, 1902 gestellt. Daraufhin beschreibt ParneS (1986) die Nautiliden aus der oberen Gevanim Formation von Makhtesh Ramon synonym als Germanonautilus salinarius Mojsisovics, 1902 und stellt alle weiteren sephardischen nautiliden Formen mit steilem Nabeleinfall gleichfalls zu Germanonautilus. Das ist eine Fehldiagnose, denn die Nabelwand von Germanonautilus fällt schräg vom Umbilikalrand zur Naht ein.
Der Gattungsbegriff Germanonautilus Mojsisovics, 1902 wird somit für die sephardische Morphe ungültig. Gleiches gilt für alle weiteren von ParneS (1986) zu Germanonautilus gestellten sephardischen Spezies.
Der steile Nabeleinfall der Steinkerne der sephardischen nautiliden Morphen ist auch ein Merkmal der Muschelkalk Gattung ?Germanonautilus. Für dieses gemeinsame Merkmal der Steinkernerhaltung wird die neue Gattung Sephardonautilus nov. gen. aufgestellt. Für dieses gemeinsame Merkmal der Steinkernerhaltung wird die neue Gattung Sephardonautilus nov. gen. aufgestellt.
Auf Grund der oben beschriebenen gemeinsamen Gat-tungsmerkmale wird die Spezies ?Germanonautilus tri-dorsatus Böttcher, 1938 in die neu aufgestellte Gattung Sephardonautilus nov. gen. transferiert und Sephardonautilus tridorsatus (Böttcher, 1938) als Typusart ge-kennzeichnet.
Sephardonautilus nov. gen.
Beschreibung
Der Windungsquerschnitt gestaltet sich von quadratisch bis trapezförmig sehr variabel. Die Flanken können glatt oder flach gewölbt sein, der schwach gerundete Mar-ginalrand ist glatt oder selten zur Lateralseite schwach wulstig-knotig skulpturiert. Die Ventralseite ist eben und kann parallel zum Marginalrand in eine schmale flache Rinne übergehen. Die Nabelweite ist variabel. Der Umbilikalrand kann bis zu einer starken Wulst ausgebildet sein. Vom Umbilikalrand fällt die Nabelwand steil zur Naht ein. Ausgehend vom letzten Septum beginnt bei 115° auf der Ventralseite des Gehäuses ein an der Basis breit gerundeter hyponomischer Sinus. Von ihm ausgehend verlaufen die Trichterränder parallel zum Marginalrand bis zur ventralen Gehäusemündung bei 165°. Die Mündung des Gehäuses biegt über den Marginalrand in einem enggerundeten Bogen zuerst konvex auf der Lateralseite nach hinten, um wenig später in einem weiten konkaven Bogen nach vorn um-zuschwenken. Dabei entstehen lateral periodisch dem konkaven Bogen folgend wulstige Schalenverdickun-gen. Die Gehäusemündung endet durch den Nabel gemessen bei 235° in einer teilweise kräftig ausgebildeten Umbilikalwulst und biegt danach senkrecht zur Naht ab. Somit erstreckt sich die Öffnung der ventralen Ge-häusemündung für den Weichkörper von 115° bis 235° über ein Bogenmaß von 120°.
Die Art Sephardonautilus tridorsatus (Böttcher, 1938) nov. comb. entstand im unteren Illyrian im Oberen Muschelkalk nach reproduktiver Isolation von der Stammart Sephardonautilus salinarius (Parnes, 1986) und starb im Unteren Fassanian im Hangenden der Gänheim-Bank aus.
Systematik
Familie Tainoeratitidae Hyatt, 1883
Gattung Sephardonautilus nov. gen.
Typusart Sephardonautilus tridorsatus (Böttcher, 1938)
Alle von PARNes (1986) als Germanonautilus beschriebenen Taxa werden in die neue Gattung Sephardonau-tilus nov. gen. gestellt:
Sephardonautilus salinarius (Parnes, 1986) nov. comb.
Sephardonautilus saharonicus (Parnes, 1986) nov. comb.
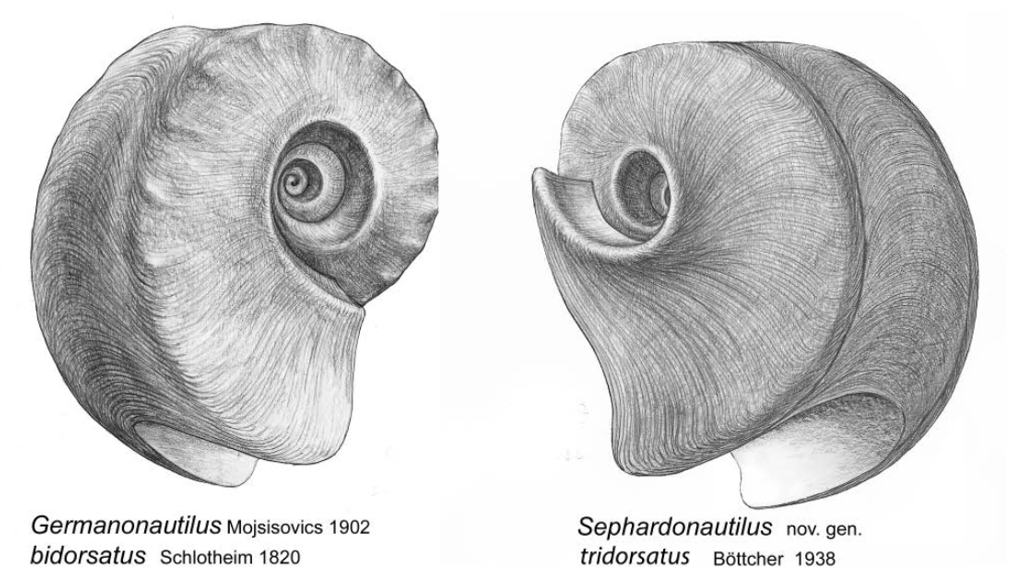
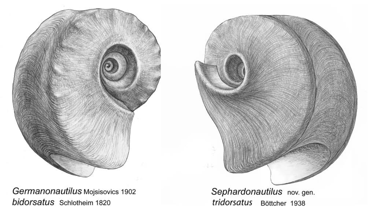
Abb. 7: Germanonautilus bidorsatus und Sephardonautilus tridorsatus
Sephardonautilus bidorsatus bidorsatus (Parnes, 1986) nov. comb.
Sephardonautilus bidorsatus jugosus (Parnes, 1986) nov. comb.
Sephardonautilus bartovi (Parnes, 1986) nov. comb.
Sephardonautilus bartovi bartovi (Parnes, 1986) nov. comb.
Sephardonautilus bartovi timidus (Parnes, 1986) nov. comb.
Sephardonautilus bartovi gravidus (Parnes, 1986) nov. comb.
Sephardonautilus ellipticus (Parnes, 1986) nov. comb.
Die morphologischen Unterschiede der Gattungen Germanonautilus und Sephardonautilus nov. gen. werden im Vergleich sichtbar. Die neue Gattung Sephardonautilus wird durch die erweiterte Beschreibung der Typusart Sephardonautilus tridorsatus (Böttcher, 1938) definiert (Rein 2016). Alle in dieser Beschreibung der Typusart aufgeführten morphologischen und physiologischen Details sind der außerordentlich guten Fossilerhaltung des thüringischen Muschelkalks zu verdanken.
Ceratites sp. der Gattung Ceratites de Haan, 1825
Der oberste Teil der Gevanim-Formation vertritt nach Parnes (1986) die Basis der trinodosus-Zone und damit das untere Illyrian. Somit entspricht das Fundniveau des Sephardonautilus salinarius chronostratigraphisch der dritten Immigrationsphase in das Muschelkalkmeer (Abb. 1c). Leider fehlen in diesem Horizont die Ammoniten. Somit gibt es aus Israel keine fossilen Belege für den ceratitiden Immigranten entsprechen. Aussagen zur Morphologie und Genealogie der unbekannten Stammform liefern zwei mäßig erhaltene Phragmokone mit kräftiger dichotom-skuptur und ceratitischer Struktur von Aref en Naqa aus dem ägyptischen Sinai. Sie wurden von Spath (1946) mit Vorbehalt als flexuose Formen zu Progonoceratites gestellt. Die Evolution der unbekannten Spezies der Gattung Ceratites de Haan endet mit der kladogenetischen Aufspaltung der Population am Ende ihres tektonisch geprägten gemeinsamen Wanderweges (grüner Ring, Abb. 1c). Durch diese allopatrische Speziation entstanden die zwei Schwester-Arten Ceratites nodosus und Ceratites tornquisti. Ein derartiger paläogeographischer und chronostratigraphischer Nachweis eines Speziationszeitpunktes ist ein evolutionsbiologisches Modellbeispiel für das biologische und evolutionäre Artkonzept in der Paläontologie (Willmann 1985, Reif 2009). Die fossil lückenlos belegte Morphokline der endemischen evolutionären Art Ceratites nodosus beginnt in der Hessisch-Thüringischen Senke (HTS) der germanischen Faunenprovinz (Abb. 2c). Die Phylogenese der Spezies Ceratites tornquisti der Gattung Ceratites de Haan beginnt in der trinodosus-Zone ohne fossilen Nachweis in einer hypothetischen vicentinisch-romanischen Faunenprovinz zwischen den Südalpen (Recoaro) und der Dobrudscha (Zibil) Abb. 2c. Durch die von außen auferlegte Separation der Populationen entstand eine reproduktive Isolation der Individuen zwischen dem endemischen Binnenmeer (Oberer Muschelkalk) und der unberechenbar offenen Palaeotethys. Die Unterbrechung des Genflusses führte zu einer ungleichmäßigen Allelen-Verteilung der getrennten Populationen und hatte eine Merkmalsdivergenz zur Folge.
4.4. Der Anisian/Ladinian Wechsel und die Immigration aus der südalpinen Faunenprovinz (Abb. 3a)
Die vierte Immigrationsphase im oberen Illyrian entspricht der Migration des Brachiopoden Punctospirella fragilis und vermutlich der Muschel Chlamys (Praechlamys) reticulata. In der südalpinen Faunenprovinz ist Punctospirella fragilis im oberen Anisian vereinzelt ohne erkennbaren Leitwert nachgewiesen (Bittner 1890, Torti & Angiolini 1997). Im Oberen Muschelkalk bildet der immigrierte Brachiopode mit der Spiriferina-Bank eine wichtige ökostragraphische Leitbank (Hagdorn & Simon 1993). Sie belegt an der Basis der evolutus-Zone ein reales Immigrations-Ereignis. Die Funde der zwei alpinen Ammoniten Flexoptychites angusto-umbilicatus und Protrachyceras recubariensis stammen jedoch aus der spinosus-Zone SW-Deutschlands. Da der Obere Muschelkalk kein zur Palaeotethys offenes Randmeer war, wirft diese erneute Immigration verschiedene Fragen auf:
– Entspricht der Fundhorizont im Muschelkalk chronostratigraphisch und biostratigraphisch wirklich dem südalpinen Horizont der Immigranten?
– Handelt es sich um passiv postmortal transportierte einzelne Schalen (Urlichs & Kurzweil 1997) oder um aktive Immigrationsversuche alpiner Ammonoideen?
Der von Urlichs & Kurzweil (1997) beschriebene Flexoptychites angusto-umbilicatus ist nach Brack & Rieber (1993) eine häufige bis zum Anisian/Ladinian-Wechsel in der secadensis-Zone ohne bedeutsame Merkmalsänderung durchgehende Spezies. Nach Urlichs (1978) wäre Protrachyceras recubariensis jedoch als problematische Spezies der curionii-Zone zu deuten, da sie morphologisch keiner bekannten Form entspricht. Dagegen stellen Brack & Rieber (1993) und Brack et al. (1995, 1996, 1999) die Morphe als Nevadites sp. in die secadensis-Zone. Mietto et al. (2018) wiederum bestimmen die Morphe als C. pemphix (Merian) aus dem Intervall zwischen der chiesense-Subzone und dem basalen Teil der curionii-Subzone.
In der südalpinen Buchenstein-Formation liegt die – GSSP – Anisian/Ladinian-Grenze am Übergang der secadensis- zur curionii-Zone (Storck et al. 2018). Aus diesem stratigraphisch bedeutsamen Grenzbereich stammen nach Ansicht der Spezialisten die Flexoptychites und Protrachyceras der südwestdeutschen spinosus-Zone. Sie besitzen deshalb einen besonderen Wertstatus für die chronostratigraphische Korrelation zwischen der germanischen und alpinen mittleren Trias. Zwei Varianten zum Immigrations-Zeitpunkt der alpinen Morphen können diskutiert werden.
Variante 1: Mit einem weiteren zeitlichen Immigrationsfenster würde damit die Anisian/Ladinian-Grenze im Oberen Muschelkalk in die mittlere spinosus-Zone verlegt und die Fundlage entspräche chronostratigraphisch und biostratigraphisch der Buchenstein-Formation. Die Fundsituation könnte kausal auf postmortal transportierte einzelne Schalen oder einen kurzzeitig erfolglosen Immigrationsversuch der alpinen Ammoniten zurückgeführt werden.
Variante 2: Bei zeitgleicher Immigration der südalpinen Ammoniten mit dem Brachiopoden Punctospirella fragilis wäre ein postmortaler Transport einzelner Schalen ausgeschlossen. Der Zeitpunkt der vierten Immigrationsphase entspräche chronostratigraphisch und biostratigraphisch einem erfolgreichen Immigrationsversuch ganzer Populationen beim Anisian/Ladinian-Wechsel. Nach erfolgter territorialer und reproduktiver Isolation entwickelten sich die immigrierten Ammoniten bis in die spinosus-Zone zu eigenen kladogenetischen Spezies. Die konservativen Flexoptychites verblieben ohne und Protrachyceras mit unbekannten morphologischen Verpunktes ist ein evolutionsbiologisches Modellbeispiel für das biologische und evolutionäre Artkonzept in der Paläontologie (Willmann 1985, Reif 2009). Die fossil lückenlos belegte Morphokline der endemischen evolutionären Art Ceratites nodosus beginnt in der Hessisch-Thüringischen Senke (HTS) der germanischen Faunenprovinz (Abb. 2c). Die Phylogenese der Spezies Ceratites tornquisti der Gattung Ceratites de Haan beginnt in der trinodosus-Zone ohne fossilen Nachweis in einer hypothetischen vicentinisch-romanischen Faunenprovinz zwischen den Südalpen (Recoaro) und der Dobrudscha (Zibil) Abb. 2c. Durch die von außen auferlegte Separation der Populationen entstand eine reproduktive Isolation der Individuen zwischen dem endemischen Binnenmeer (Oberer Muschelkalk) und der unberechenbar offenen Palaeotethys. Die Unterbrechung des Genflusses führte zu einer ungleichmäßigen Allelen-Verteilung der getrennten Populationen und hatte eine Merkmalsdivergenz zur Folge.
4.4. Der Anisian/Ladinian Wechsel und die Immigration aus der südalpinen Faunenprovinz (Abb. 3a)
Die vierte Immigrationsphase im oberen Illyrian entspricht der Migration des Brachiopoden Punctospirella fragilis und vermutlich der Muschel Chlamys (Praechlamys) reticulata. In der südalpinen Faunenprovinz ist Punctospirella fragilis im oberen Anisian vereinzelt ohne erkennbaren Leitwert nachgewiesen (Bittner 1890, Torti & Angiolini 1997). Im Oberen Muschelkalk bildet der immigrierte Brachiopode mit der Spiriferina-Bank eine wichtige ökostragraphische Leitbank (Hagdorn & Simon 1993). Sie belegt an der Basis der evolutus-Zone ein reales Immigrations-Ereignis. Die Funde der zwei alpinen Ammoniten Flexoptychites angusto-umbilicatus und Protrachyceras recubariensis stammen jedoch aus der spinosus-Zone SW-Deutschlands. Da der Obere Muschelkalk kein zur Palaeotethys offenes Randmeer war, wirft diese erneute Immigration verschiedene Fragen auf:
– Entspricht der Fundhorizont im Muschelkalk chronostratigraphisch und biostratigraphisch wirklich dem südalpinen Horizont der Immigranten?
– Handelt es sich um passiv postmortal transportierte einzelne Schalen (Urlichs & Kurzweil 1997) oder um aktive Immigrationsversuche alpiner Ammonoideen?
Der von Urlichs & Kurzweil (1997) beschriebene Flexoptychites angusto-umbilicatus ist nach Brack & Rieber (1993) eine häufige bis zum Anisian/Ladinian-Wechsel in der secadensis-Zone ohne bedeutsame Merkmalsänderung durchgehende Spezies. Nach Urlichs (1978) wäre Protrachyceras recubariensis jedoch als problematische Spezies der curionii-Zone zu deuten, da sie morphologisch keiner bekannten Form entspricht. Dagegen stellen Brack & Rieber (1993) und Brack et al. (1995, 1996, 1999) die Morphe als Nevadites sp. in die secadensis-Zone. Mietto et al. (2018) wiederum bestimmen die Morphe als C. pemphix (Merian) aus dem Intervall zwischen der chiesense-Subzone und dem basalen Teil der curionii-Subzone.
In der südalpinen Buchenstein-Formation liegt die – GSSP – Anisian/Ladinian-Grenze am Übergang der secadensis- zur curionii-Zone (Storck et al. 2018). Aus diesem stratigraphisch bedeutsamen Grenzbereich stammen nach Ansicht der Spezialisten die Flexoptychites und Protrachyceras der südwestdeutschen spinosus-Zone. Sie besitzen deshalb einen besonderen Wertstatus für die chronostratigraphische Korrelation zwischen der germanischen und alpinen mittleren Trias. Zwei Varianten zum Immigrations-Zeitpunkt der alpinen Morphen können diskutiert werden.
Variante 1: Mit einem weiteren zeitlichen Immigrationsfenster würde damit die Anisian/Ladinian-Grenze im Oberen Muschelkalk in die mittlere spinosus-Zone verlegt und die Fundlage entspräche chronostratigraphisch und biostratigraphisch der Buchenstein-Formation. Die Fundsituation könnte kausal auf postmortal transportierte einzelne Schalen oder einen kurzzeitig erfolglosen Immigrationsversuch der alpinen Ammoniten zurückgeführt werden.
Variante 2: Bei zeitgleicher Immigration der südalpinen Ammoniten mit dem Brachiopoden Punctospirella fragilis wäre ein postmortaler Transport einzelner Schalen ausgeschlossen. Der Zeitpunkt der vierten Immigrationsphase entspräche chronostratigraphisch und biostratigraphisch einem erfolgreichen Immigrationsversuch ganzer Populationen beim Anisian/Ladinian-Wechsel. Nach erfolgter territorialer und reproduktiver Isolation entwickelten sich die immigrierten Ammoniten bis in die spinosus-Zone zu eigenen kladogenetischen Spezies. Die konservativen Flexoptychites verblieben ohne und Protrachyceras mit unbekannten morphologischen Verletzung auf dem Venter löste Spath (1934) den Fund aus der Gattung Ceratites de Haan 1825 und stellte ihn unberechtigt in eine eigene Gattung Alloceratites (eine Verletzung bedingt keine taxonomische Änderung).
Abb. 6: Cerattes schmiat und Neoclypues . peregrinus
a) Ceratites schmidi, Original Philppi 1901. Abb. 5 und Sa. aa) Ceratites schmidi, Orginal Philipp 1901. Abb. Sb und Sc. b) Ceratites schmidi Gipsabgusse vom zweiten rund 196l (palaont. Sammlung Halle Nr. H 61). Museum TU Freiberg Nr. 123/1, C) Ceratites schmidi, Original vom dritter Fund 1970, paläont. Sammlung TU Freiberg Nr. 144/1, d) Neoclypites ? peregrinus, Original, paläont. Sammlung TU Freiberg Nr. 144/2
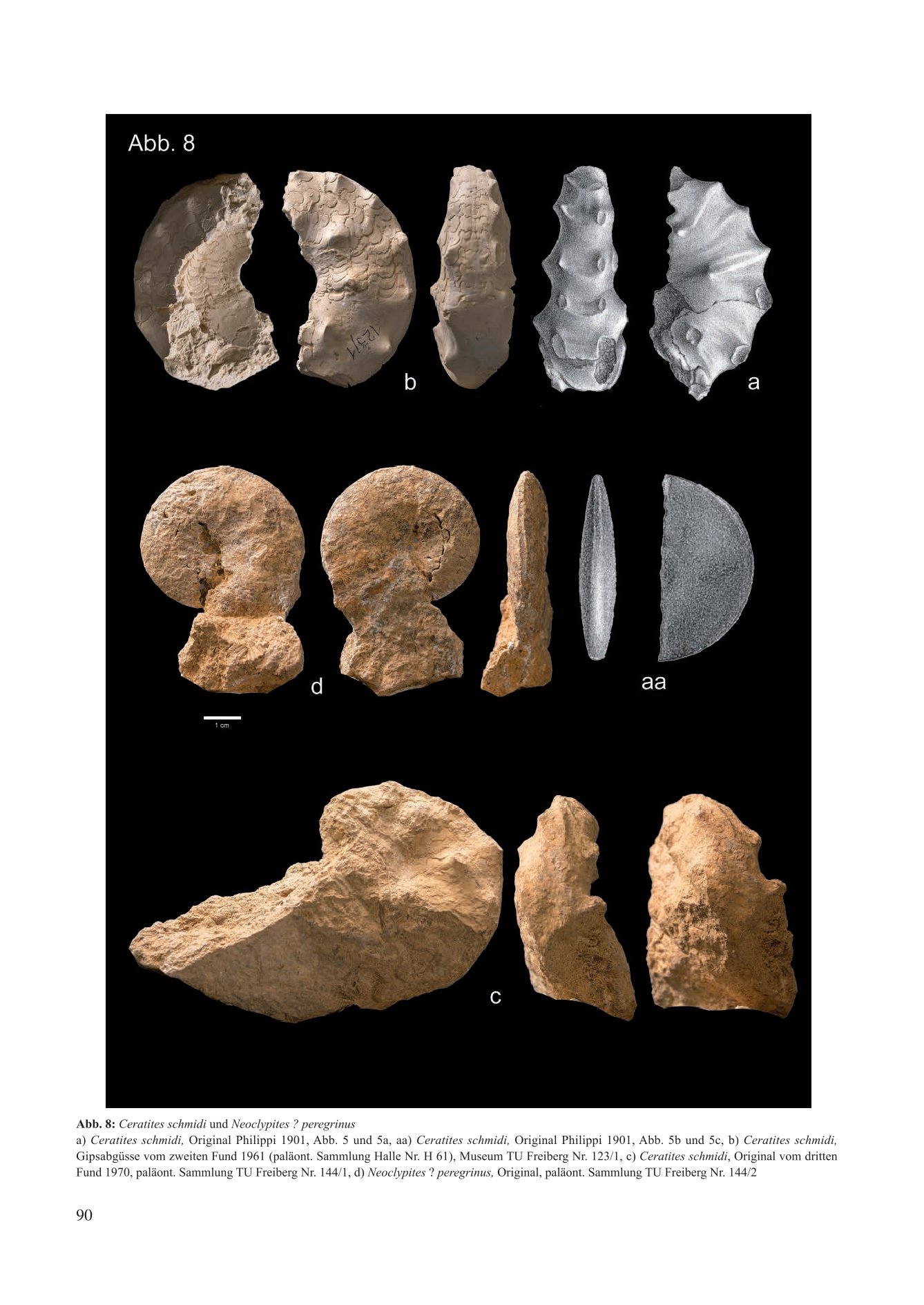
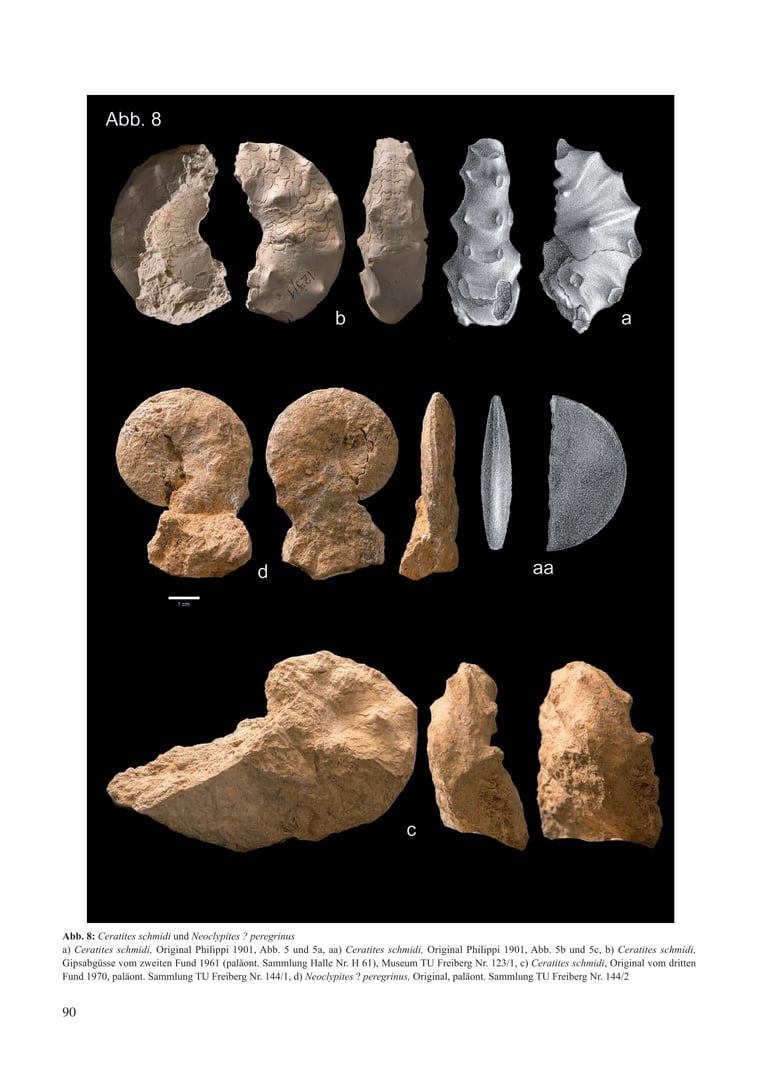
Mit dem Fund eines zweiten Ceratiten aus dem thüringischen Grenzdolomit in der Ziegeleigrube Reisdorf bei Apolda (1961) ergab sich ein taxonomisches Problem, denn das neue Belegstück besitzt eine skulpturfreie glatte Externseite (Abb. 8 c/d). Um für den Reisdorfer Fund nicht eine neue Art und eine neue Untergattung schaffen zu müssen, stellte Müller (1969) daraufhin Ceratites schmidti wieder zur Gattung Ceratites de Haan und versetzte das Taxon Alloceratites Spath, 1934 in den Rang einer Untergattung.
Mit weiteren Neufunden aus dem Grenzdolomit der Ziegeleigrube Reisdorf (Abb. 8 e/f) veränderte sich die taxonomische Zuordnung erneut (Müller 1973). Beim ersten Belegstück handelt es sich um die „Normalform“ eines Ceratites schmidti. Das zweite hat ein flaches scheibenförmiges Gehäuse mit einer teilweise median auf dem Venter verlaufenden Furche. Es wird als neue Spezies Neoclypites ? peregrinus n. sp. beschrieben. Kozur (1971) nimmt an, dass Israelites ramonensis aus dem Ladinian Israels gewisse übereinstimmende Merkmale der beiden Morphen besitzt und vermutet, chronostratigraphisch bedingt, engere verwandtschaftliche Beziehungen zwischen Alloceratites und Hungarites als mit Ceratites. So löst Müller (1973) Ceratites schmidti endgültig von der Gattung Ceratites de Haan und stellt die Ceratiten aus dem Thüringer Grenzdolomit allein zu Alloceratites Spath, 1934.
Mit der kladogenetischen Aufspaltung einer Population der Gattung Ceratites de Haan in die Schwesterarten Ceratites tornquisti und Ceratites nodosus klärt sich der genealogische Ursprung (Abb. 2c). Die endemische Phylogenese der evolutionären Art Ceratites nodosus endet im Oberen Muschelkalk an der Basis der Erfurt-Formation mit dem Aussterben der Spezies. Parallel verläuft die heterogene Evolution der Schwesterart räumlich und reproduktiv getrennt in der Palaeotethys. Unabhängig von weiteren Artaufspaltungen in der Phylogenese verbleiben alle Nachfahren der Art Ceratites tornquisti Teil der Gattung Ceratites de Haan. Deshalb erfolgte die Immigration im Longobardian aus einem unbekannten Refugium der Palaeotethys als Spezies Ceratites schmidti (Abb. 3c).
Unsicher ist lediglich die Deutung der medianen Rinne des „Neoclypites ? peregrinus“. Das Gehäuse steckte ursprünglich auf der Ventralseite in einem körnigen dolomitischen Block. Nur wenn die Rinne mechanisch durch Herausbrechen entstand, wäre es ein juveniles Individuum der Gattung Ceratites de Haan und ein Nachfahre der ursprünglichen Schwesterart Ceratites tornquisti. Ansonsten bleibt die Genealogie der Spezies Neoclypites ? peregrinus unbekannt.
Ein anderes Problem bereitet die Migration des Germanonautilus jugatonodosus in den Grenzdolomit. Nach Merki (1961) endete der Obere Muschelkalk im östlichen Schweizer Jura mit dem Äquivalent des Grundgipses. Demnach wäre das südliche Randgebiet des Muschelkalkmeeres in der nördlichen Schweiz zu einem Rückzugsraum der marinen Muschelkalk-Fauna auch für Germanonautilus bidorsatus geworden (Rein 2014). Nachdem Pietsch et al. (2016) die stratigraphische Reichweite des Unteren Keupers korrigierten, muss die Immigration des Germanonautilus jugatonodosus, ohne Bezug zur Phylogenese des Germanonautilus bidorsatus, revidiert werden (Urlichs 2000, 2015, Rein 2015).
8. Fazit
Als neu entstandenes Ökosystem ist der Obere Muschelkalk mit seinen stets kurzzeitig wechselnden marinen Verbindungen zu verschiedenen Faunen-Provinzen der Palaeotethys und Neotethys ein tektonisch dominiertes Binnenmeer. Immigrierte Makrofossilien ermöglichen die biostratigraphische und chronostratigraphische Korrelation und Rekonstruktion der von Germanonautilus bidorsatus und Sephardonautilus tridorsatus chronologisch markierten hypothetischen Wanderwege mit hypothetischen Pforten in das Binnenmeer.
Als erster migrierte in der unteren trinodosus-Zone Germanonautilus salinarius aus der austroalpin-dinarischen Faunenprovinz der Palaeotethys über eine hypothetische helvetisch-alemannische Pforte bis in das Beckeninnere. Der aus der Stammart kladogenetisch entstandenen Spezies Germanonautilus bidorsatus gelang die physiologische Anpassung an alle wechselnden Ökosysteme von der Diemel-Formation bis zur Erfurt-Formation.
Aus der austroalpinen Faunenprovinz migrierte in der mittleren trinodosus-Zone Sturia sansovinii auf dem gleichen Weg bis in das Beckeninnere und bildete kladogenetisch die neue Spezies Sturia brandti n. sp.
Der bedeutendste Schritt der Besiedlung des Ökosystems Oberer Muschelkalk erfolgte im unteren Illyrian aus der Sephardischen Faunenprovinz. Durch eine einmalig kurzzeitig tektonisch geschaffene burgundisch-rheinische Pforte migrierten fremdartige Cephalopoden aus der Neotethys in das Binnenmeer. Mit Sephardonautilus tridorsatus nov. comb. als Typusart der neuen Gattung Sephardonautilus und der Stammart Sephardonautilus salinarius wird die verwandtschaftliche Verbundenheit zwischen den Individuen des Muschelkalkmeeres und der sephardischen Faunenprovinz dokumentiert.
Gleiches gilt für die nach allopatrischer Speziation als Schwesterart aus einer unbekannten Spezies der Gattung Ceratites de Haan der Sephardischen Faunenprovinz entstandene evolutionäre Spezies Ceratites nodosus. Die Stammart Sephardonautilus salinarius aus der trinodosus-Zone der oberen Gevanim-Formation von Makhtesh Ramon belegt die chronologische Korrelation des Migrationsweges des kladogenetisch entstandenen Sephardonautilus tridorsatus im Oberen Muschelkalk. Die Morphokline der beispiellos eurypotenten evolutionären Art Ceratites nodosus ermöglicht die lückenlose biostratigraphische Gliederung des Oberen Muschelkalkes.
Mit der chronologischen Korrelation der Stammarten von Germanonautilus bidorsatus und Sephardonautilus tridorsatus zu den austroalpinen und sephardischen Faunenprovinzen wird die trinodosus-Zone die stratigraphische Basis des Oberen Muschelkalkes.
Alle immigrierten alpinen Ammoniten erweisen sich beim Vergleich zur eurypotenten Spezies Ceratites nodosus als physiologisch nur bedingt anpassungsfähig. Die Immigranten Pinacoceras und Gymnites aus der unterladinischen Schreyeralm-Formation schaffen den Weg bis in das Beckenzentrum. Das physiologische Integrationsvermögen der kladogenetisch entstandenen Spezies Parapianoceras thiemei und Gymnites bruneli bleibt jedoch begrenzt.
Überraschend erscheint das Anpassungsverhalten der südalpinen Morphen Flexoptychites angusto-umbilicatus und Protrachyceras recubariensis. Sie als passiv transportierte isolierte Schalenreste in der spinosus-Zone SW-Deutschlands zu betrachten fällt aus verschiedenen Gründen schwer. Favorisiert wird eine zweite aktive Variante. Bei zeitgleicher Immigration mit dem Brachiopoden Punctospirella fragilis wird ein postmortaler Transport einzelner Schalen ausgeschlossen. Der Zeitpunkt der vierten Immigrationsphase entspräche chronostratigraphisch und biostratigraphisch einem erfolgreichen Immigrationsversuch ganzer Populationen beim Anisian/Ladinian-Wechsel. Die immigrierten Flexoptychites und Protrachyceras überleben im südwestdeutschen Raum mit physiologischer Anpassung bis in die spinosus-Zone. In dieser Zeit erwerben sie kladogenetisch einen neuen Artstatus.
Für die Festlegung des Anisian/Ladinian-Wechsels ergäbe sich die Spiriferina-Bank mit dem Wechsel von der compressus-Zone zur evolutus-Zone. Er korreliert in der südalpinen Faunenprovinz mit dem Wechsel von der secadensis-Zone zur curionii-Zone.
Die Evolution der unbekannten Spezies der Gattung Ceratites de Haan aus der Sephardischen Faunenprovinz endet mit der allopatrischen Aufspaltung in die zwei Schwester-Arten Ceratites nodosus und Ceratites tornquisti. Die Phylogenese der Spezies Ceratites tornquisti beginnt im unteren Illyrian in der Palaeotethys und endet im Longobardian im Grenzdolomit in Thüringen als Ceratites schmidti. Aufgrund seiner fastigaten Skulpturbildungen wurde C. schmidti von Spath (1934) aus der Gattung Ceratites de Haan gelöst und unberechtigt in eine eigene Gattung Alloceratites gestellt. Die traumatisch bedingte morphologische Änderung eines Individuums verändert keinen Gattungs-Status. Mit der Klärung des genealogischen Ursprungs von Ceratites tornquisti, der Schwesterart von Ceratites nodosus, verbleiben alle Nachfahren der Art Teil der Gattung Ceratites de Haan. Die Klärung der bislang unbekannten Phylogenese der Spezies Ceratites tornquisti in den Lebensräumen Dobrudscha, Sardinien, Vicentin, Provence und Thüringen bleibt einer detaillierten Analyse vorbehalten.
Dank
Bei Herrn M. Hartmann, Naturkundemuseum Erfurt, bedanke ich mich für die vielseitige Unterstützung meiner Arbeit, bei Herrn Prof. Dr. F. Hirsch (Uni Naruto) bedanke ich mich für fachlichen Rat, Herrn S. Brandt (Kornhochheim) danke ich für die Fotoarbeiten, Herrn Dr. J. A. Perez Valera (Uni Madrid) danke ich für die Hilfe bei der Beschaffung wichtiger Literatur und bei Herrn Prof. Dr. J. W. Schneider (TU Freiberg) bedanke ich mich für die Ausleihe der Originale der Ceratiten aus dem Grenzdolomit.
Literatur
Adams, A. & L. W. Diamond (2019): Facies and depositional environments of the Upper Muschelkalk (Schinznach Formation, Middle Triassic) in northern Switzerland. – Swiss Journal of Geosciences, 25 S., 14 Abb.
Anastasiu, V. (1898): Contribution ix l’etude geologique de la Dobrogea. Terrains secondaires. – These pres. a la faculte des Sciences de Paris, p. 46.
Bachmann, G. H., M. Geluk, J. Warrington, A. Becker, G. Beutler, H. Hagdorn, M. Hounslow, E. Nitsch, H.-G. Röhl, Th. Simon & A. Szulc (2010): with contributions of Dusar, M., Nielsen, L. H., Barnasch, J., Franz, M.: Triassic. – In: Doornenbal, H. & A. Stevenson (eds.): Petroleum Geological Atlas of the Southern Permian Basin Area, 149–173; Houten/The Netherlands (EAGE).
Bittner, A. (1890): Brachiopoden der alpinen Trias. – Abhandlungen der kaiserlich-königlichen geologischen Reichsanstalt XIV, 325 S., 41 Taf.
Böttcher, R. J. (1938): Versteinerungen des Oberen Muschelkalks bei Ohrdruf als aufschlussreiche Dokumente für die Geschichte des deutschen Muschelkalkmeeres. – Beiträge zur Geologie von Thüringen 5: 99–105.
Brack, P. & H. Rieber (1993): Towards a better definition of the Anisian/Ladinian boundary: New biostratigraphic data and correlations of boundary sections from the Southern Alps. – Eclogae geologicae Helvetiae 86/2: 415–527, 17 Abb., 14 Taf.
Brack, P., H. Rieber & M. Urlichs (1999): Pelagic successions in the Southern Alps and their correlation with the Germanic Middle Triassic. – Zentralblatt Geologie und Paläontologie Teil I, Heft 7–8, 853–976, 8 Fig., Stuttgart.
Brack, P., H. Rieber & R. Mundil (1995): The Anisian/Ladinian Boundary interval at Bagolino (Southern Alps, Italy): In. Summary and news results on ammonoid horizons and radiometric age dating. – Albertiana 15: 45–56, 3 Fig.
Brack, P., H. Rieber & M. Urlichs (1996): The new „high resolution Middle Triassic ammonoid standard scale“ proposed by Triassic researchers from Padova – a discussion of the Anisian/Ladinian boundary interval. – Albertiana 17: 42–50.
Brandt, S. & S. Rein (2019): Die Chronospezies Ceratites flexuosus von Geilsdorf – Ein evolutionsbiologisches Modellfall der evolutionären Art Ceratites nodosus. – VERNATE 38: 53–76.
Broili, F. (1927): Eine Muschelkalkfauna aus der Nähe von Saalfelden. – Sitzungsberichte der mathematisch-naturwissenschaftlichen Abteilung der BAK, Heft III: 229–242.
Diener, C. (1900): Die triadische Cephalopodenfauna der Schichlinghöhe bei Hallstatt. – in: Beiträge zur Paläontologie und Geologie Österreich-Ungarns und des Orients; Ed. Dr. G. v. Arthaber; Band XIII, Wien und Leipzig 1901; Seite 3–42 und 3 Tafeln.
Diener, C. (1916): Einige Bemerkungen zur Nomenklatur der Triascephalopoden. – Centralblatt für Mineralogie, Geologie und Paläontologie 1916: 97–105.
De Haan, G. (1825): Monographiae Ammoniteorum et Goniatiteorum Specimen. – II + 168 S.; Leyden (Hazenberg).
Hagdorn, H. & T. Simon (1993): Ökostragraphische Leitbänke im Oberen Muschelkalk. – In: Hagdorn, H. & A. Seilacher (eds.): Muschelkalk. Schöntaler Symposium 1991, Stuttgart, Korb (Goldschneck); 193–208, 15 Abb.
Hauer, F. v. (1887): Die Cephalopoden des Bosnischen Muschelkalkes von Han Bulog bei Sarajevo. – Denkschrift der mathematisch-naturwissenschaftlichen Classe der kaiserlichen Akademie der Wissenschaften, 8 Taf.
Hauer, F. v. (1892): Cephalopoden aus der Trias von Bosnien I. – Denkschrift der mathematisch-naturwissenschaftlichen Classe der kaiserlichen Akademie der Wissenschaften, 15 Taf.
Hauer, F. v. (1896): Cephalopoden aus der Trias von Bosnien II. – Denkschrift der mathematisch-naturwissenschaftlichen Classe der kaiserlichen Akademie der Wissenschaften, 13 Taf.
Hohenstein, V. (1913): Beiträge zur Kenntnis des Mittleren Muschelkalks und des unteren Trochitenkalks am östlichen Schwarzwaldrand. – Geologisch-paläontologische Abhandlungen (16), N.F.12: 175–272, 12 Abb., Taf. 12–19.
Knaust, D. (1991): Ein bemerkenswerter Nautiliden-Kiefer (Rhyncholites hirundo Faure-Biguet, Cephalopoda) aus dem Mittleren Muschelkalk (Oberanis) der Germanischen Trias. – Veröffentlichungen des Naturkundemuseums Erfurt 10: 58–65, 4 Abb., 1 Taf.
Kozur, H. (1971): Mikropaläontologie, Biostratigraphie und Biofazies der germanischen Mitteltrias. – Freiberg Bergakademie, Dissertation, Freiberg.
Kozur, H. (1974): Biostratigraphie der germanischen Mitteltrias, Teil I und II mit Anlagen. – Freiberg Forschungshefte C, 280, I: 1–56, II: 1–71.
Kummel, B. (1960): Middle Triassic Nautiloids from Sinai, Egypt and Israel. – Bulletin of the Museum of Comparative Zoology at Harvard College 123 (7): 291–304, 2 Pls.
Merki, P. J. (1961): Der Obere Muschelkalk im östlichen Schweizer Jura. – Birkhäuser AG, 263 S., Basel.
Mietto, P., S. Manfrin & M. Rigo (2018): Middle Triassic ammonoid fauna from the Recoaro and Tretto areas (NE Italy) and its stratigraphic and paleobiogeographic evidence. – Bollettino della Società Paleontologica Italiana 53 (3), 217–250, 8 Fig.
Mojsisovics, E. v. (1873–1902): Das Gebirge um Hallstatt; Abt. I, Die Cephalopoden der Hallstätter Kalke. – Abhandlungen der kaiserlich-königlichen Geologischen Reichsanstalt 6: 1–356, pl. 1–94.
Mojsisovics, E. v. (1882): Cephalopoden der mediterranen Triasprovinz. – Abhandlungen der kaiserlich-königlichen Geologischen Reichsanstalt 10: 1–322, 1–94.
Mojsisovics, E. v. (1902): Das Gebirge um Hallstatt. – Abhandlungen der kaiserlich-königlichen Geologischen Reichsanstalt, 3. Lief. (Supplement): 175–356, Taf. 1–23.
Müller, A. H. (1969): Ein Ceratit (Ceratites cf. schmidti, Ammonoidea) aus dem Unterkeuper (Grenzdolomit) des Germanischen Triasbeckens. – Monatsberichte der Deutschen Akademie der Wissenschaften zu Berlin 11, 2: 122–132, 5 Abb., 2 Taf.
Müller, A. H. (1970): Neue Funde seltener Ceratiten aus dem germanischen Muschelkalk und Keuper. – Monatsberichte der Deutschen Akademie der Wissenschaften zu Berlin 12, 8: 632–642, 16 Abb., 1 Taf.
Müller, A. H. (1973): Über Ammonoidea (Cephalopoda) aus dem Grenzdolomit des germanischen Unterkeupers. – Zeitschrift für geologische Wissenschaft 1, 8: 935–945, 9 Abb., 1 Taf.
Pietsch, J. S., Wetzel, A. Jordan, P. (2016): A new lithostratigraphic scheme for the Schinznach Formation (upper part of the Muschelkalk Group of northern Switzerland). – Swiss Journal of Geosciences: 285–307, 15 Fig.
Reif, W. E. (2009): Artabgrenzung und das Konzept der evolutiven Art in der Paläontologie. – Journal of Zoological Systematics and Evolutionary Research 22 (3): 263–286, 2 Abb.
Rein, S. (2007): Die Biologie der Ceratiten der flexuosus-, sequens und pulcher und semipartitus/meissnerianus-Zone. – Entstehung und Aussterben der Biospezies Ceratites nodosus. – Veröffentlichungen des Naturkundemuseums Erfurt 26: 39–67, 32 Abb., 6 Taf., 3 Prof.
Rein, S. (2008): Ceratites avitus auf Mallorca? – Die Muschelkalkammonoideen des Museu Balear de Sciencies Naturals Sóller. – VERNATE 27: 5–19, 7 Abb.
Rein, S. (2009): Stratigraphie und Fossilführung des Oberen Muschelkalkes bei Eisenach und Jena. – VERNATE 28: 31–49, 10 Abb., 5 Taf.
Rein, S. (2014): Germanonautilus im Unteren Keuper (Trias, Erfurt-Formation) – von Trematodiscus jugatonodosus bis Germanonautilus bidorsatus. – Semana 29: 37–44, 11 Abb.
Rein, S. (2015): Erwiderung auf Urlichs vorstehende „Entgegnung zu Rein (2014)“: Germanonautilus im Unteren Keuper (Trias, Erfurt-Formation) – von Trematodiscus jugatonodosus bis Germanonautilus bidorsatus. – Semana 30: 57–58, 1 Abb.
Rein, S. (2016): Germanonautilus bidorsatus und ?Germanonautilus tridorsatus (Nautiloidea) aus dem Oberen Muschelkalk (Mitteltrias, Anis–Ladin). Teil 1: Speziation, Ontogenese und Phylogenese. – VERNATE 35: 31–65, 13 Abb., 11 Taf.
Rein, S. (2017): Speziation, Phylogenie und Ontogenie der Biospezies Ceratites nodosus und das Chronospezies Konzept. – VERNATE 36: 47–64, 18 Abb.
Rein, S. & R. Werneburg (2010): Parapianoceras und Gymnites (Ammonoidea) aus der enodis/posseckeri-Zone im Oberen Muschelkalk (Mittel-Trias, Ladin) Thüringens. – Semana 25: 87–100, 9 Abb., 1 Taf.
Seidel, G. (2013): Stratigraphie, Fazies und geologische Stellung des Zechsteins und der Trias Thüringens. – Beiträge zur Geologie von Thüringen N.F. 20: 21–78, 27 Abb., 12 Tab.
Schlotheim (1820): Petrefaktenkunde. – Gotha.
Schmidt, M. (1928): Die Lebewelt unserer Trias. – Hohenloh’sche Buchhandlung F. Rau; 461 S., 2300 Abb., Öhringen.
Schröder, W. (1977): Ein besonderer Fund … – Der Aufschluss 28, 110, 1 Abb.
Stampfli, G. M. & G. D. Borel (2002): A plate tectonic model for the Paleozoic and Mesozoic constrained by dynamic plate boundaries and restored synthetic oceanic isochrones. – Earth and Planetary Science Letters 196: 17–33.
Schnetzer, R. (1934): Die Muschelkalkfauna des Ofenbachgrabens bei Saalfelden. – Palaeontographica 81: 1–160, 15 Textabb., Taf. 1–6.
Spath, L. F. (1934): The Ammonoidea of the Trias (II). – Catalogue of Fossils, Ceph. Brit. Mus. IV. 521 S., 18 Taf.
Spath, L. F. (1946): The Middle Triassic Cephalopoda from Sinai. – Institute Egypt Bulletin 27: 425–426, pl. III.
Storck, J.-C., P. Brack, J.-F. Wotzlaw & P. Ulmer (2018): Timing and evolution of Middle Triassic magmatism in the Southern Alps (northern Italy). – Journal of the Geological Society; 16 S., 12 Fig., 1 Tab.
Szulc, J. (2000): Middle Triassic evolution of the northern Peri-Tethys area as influenced by early opening of the Tethys Ocean. – Annales Societatis Geologorum Poloniae 70: 1–48.
Tollmann, A. (1976): Analysen des klassischen nordalpinen Mesozoikums. – Franz Deuticke Wien, 581 S.
Tornquist, A. (1898): Neue Beiträge zur Geologie und Paläontologie der Umgebung von Recoaro und Schio (im Vicentin). 2. Beitrag: Die Subnodusus-Schichten. – Zeitschrift der deutschen Geologischen Gesellschaft 50: 635–694, Taf. XX–XXIII.
Tornquist, A. (1901): Das Vorkommen von nodosen Ceratiten auf Sardinien und über die Beziehungen der mediterranen zu den deutschen Nodosen. – Centralblatt Mineralogie etc.: 385–396
Weitere Artikel
Kontaktieren Sie uns.
+49 176 61287877
brabast@gmail.com
gesponsert von: