

Ein weiterer Einblick in die terrestrische Fortbewegung des Perms: Eudibamus cursoris
Dieser Artikel beschreibt die wissenschaftliche Rekonstruktion von Eudibamus cursoris durch Sebastian Brandt – eines 280 Millionen Jahre alten permzeitlichen Landwirbeltiers aus der Fossilfundstelle Bromacker in Thüringen. Es zeigt Eudibamus als den bislang frühesten bekannten zweibeinigen Läufer und rekonstruiert anhand detaillierter Fossilanalysen seine Anatomie, Fortbewegung, Hautstruktur und Ökologie. Grundlage dafür sind innovative Modellbautechniken, die sich konsequent an einem aktualistischen Ansatz der aktuellen paläontologischen Forschung orientieren.
Sebastian Brandt


Einführung
Eudibamus ist eine ausgestorbene Gattung aus der Familie der Bolosauridae, die sich während des Perms vor etwa 280 Millionen Jahren entwickelte. Diese rätselhaften Wirbeltiere wurden den Parareptilia („reptilienähnlich“) zugeordnet, da sie trotz ihres höheren Alters reptiliomorphe anatomische Merkmale aufweisen und damit bekannten Reptilien zeitlich vorausgehen. Die Parareptilia stellen ein einzigartiges evolutionäres Experiment dar, das noch vor der Aufspaltung der Wirbeltiere in Amphibien, Reptilien und Säugetiere entstand.
Das einzige bekannte Skelett von Eudibamus cursoris wurde im renommierten Bromacker-Steinbruch in Thüringen (Mitteldeutschland) entdeckt, einer der weltweit bedeutendsten Fossilfundstellen für frühe landlebende Wirbeltiere.
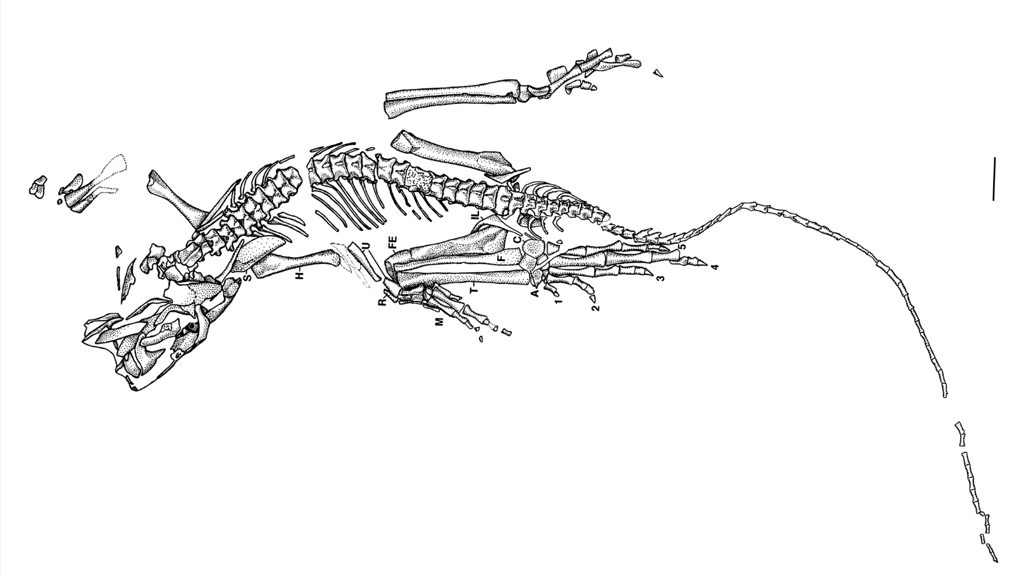
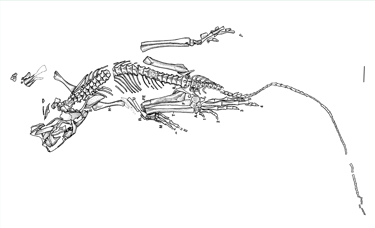
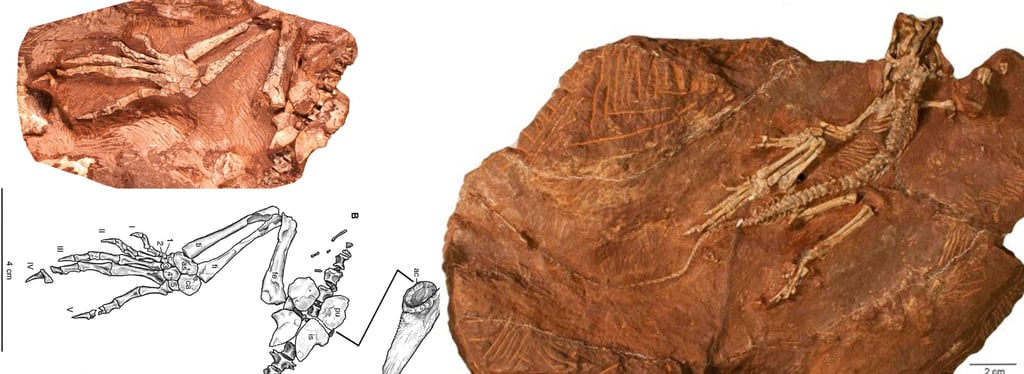

Die Bromacker-Funde belegen ein komplexes terrestrisches Ökosystem des mittleren Perms mit einer vielfältigen Fauna und Flora, darunter frühe landlebende Wirbeltiere wie Orobates, Diadectes, Dimetrodon und weitere Arten. Die Sand- und Tonsteinsedimente der Fundstelle entstanden auf einem kontinentalen Hochplateau, das von periodischen Wasserläufen, Tümpeln und Überflutungsebenen geprägt war.
Diese permzeitliche Region lag weit entfernt von marinen Einflüssen und war einem extremen kontinentalen Klima mit starken täglichen Temperaturschwankungen ausgesetzt. Sie existierte zur Zeit des Superkontinents Pangäa, noch bevor dieser in die heutigen Landmassen zerfiel. Nachgewiesene periodische Temperaturen unter 0 °C zeigen, dass sich Fauna und Flora spezielle anatomische Anpassungen entwickelten, um diese klimatischen Extreme zu überstehen. Die Wirbeltierfauna dieser Zeit und Region, als „Ursaurier“ bezeichnet, stellt zwar keine direkten Vorfahren der später auftretenden Dinosaurier dar, repräsentiert jedoch die ersten eindeutig belegten, vollständig terrestrischen Wirbeltiere der Evolutionsgeschichte.


Fakten für die Rekonstruktion
Der Holotyp von Eudibamus cursoris (MNG 8852, Stiftung Schloss Friedenstein Gotha, Deutschland) wurde 1993 entdeckt. Er besteht aus einem nahezu vollständigen, artikulierten Skelett, das bis auf einen leicht disartikulierten und diagenetisch veränderten Schädel außergewöhnlich gut erhalten ist. Ein weiteres Exemplar (MNG 12895) umfasst ein gut erhaltenes Becken sowie ein artikuliertes Hinterbein. Sämtliche anatomischen Maße dieser Funde wurden im Maßstab 1:1 in das Rekonstruktionsmodell übertragen.
Die wesentlichen anatomischen Merkmale von Eudibamus cursoris sind:
– Kleine, grazile und sehr bewegliche Körpergestalt
– Langer, schlanker Rumpf mit kurzen Rippen und röhrenförmigem Brustkorb
– Grazile Vorderextremitäten mit langen Fingern und kleinen Krallen
– Lange, schlanke Hinterextremitäten mit verlängerten Unterschenkeln und nur schwach bekrallten Zehen
– Füße mit angedeuteter Fersenstruktur
– Kurzer, kräftiger Hals ohne Halsrippen
– Dreieckiger Schädel mit großen Augenhöhlen
– Spezialisierte Bezahnung mit stiftförmigen Vorderzähnen und konischen Hinterzähnen
– Schlanke Kieferknochen
– Langer, graziler Schwanz mit kräftigen proximalen Rippen
– Sehr kleines Becken mit diagonal ausgerichteten Hüftgelenken
– Deutliche skelettmorphologische Parallelen zum rezenten Physignathus cocincinus
Die Skelettmorphologie von Eudibamus weist insgesamt auf einen extrem schlanken Körperbau hin, der eine hohe Beweglichkeit und spezialisierte terrestrische Fortbewegung nahelegt.
Die vorliegenden Rekonstruktionsmodelle integrieren ökologische und biologische Erkenntnisse, die im Rahmen intensiver Recherche sowie interdisziplinärer Zusammenarbeit mit Paläontologen wie Dr. Thomas Martens, Dr. Tom Hübner, Dr. Ralf Werneburg und weiteren Fachwissenschaftlern erarbeitet wurden.
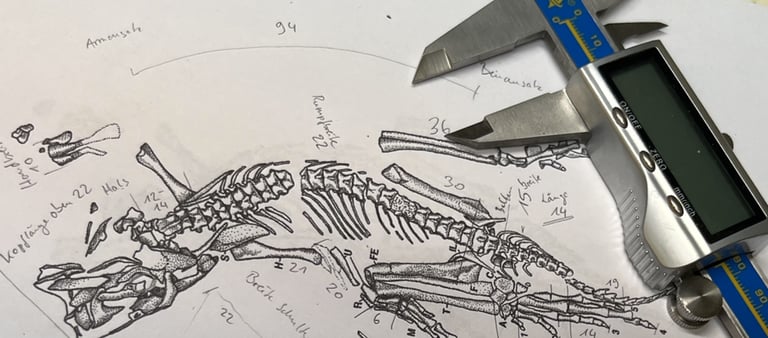
Rekonstruktion der funktionellen Morphologie
Fortbewegung
Berman et al. (2000) identifizierten Eudibamus cursoris als den bislang frühesten bekannten Wirbeltiervertreter, der zu zweibeiniger Fortbewegung fähig war. Für die vorliegende Arbeit wurden zwei Rekonstruktionsmodelle entwickelt: eines in quadrupeder Haltung, die als primäre Fortbewegungsform interpretiert wird, und ein weiteres, das einen etwas unbeholfenen zweibeinigen Lauf darstellt. Die Analyse der Lokomotion bezog Bewegungsstudien rezenter Arten ein, insbesondere die ungewöhnlichen zweibeinigen Laufbewegungen von Basiliscus plumifron, um die funktionelle Morphologie eines Wechselns der Fortbewegungsweise besser zu verstehen.
Die hauptsächliche Fortbewegungsart von Eudibamus war quadruped. Das Tier bewegte sich vermutlich entlang schlammiger Uferzonen von Tümpeln sowie durch Pioniervegetation der Bromacker-Landschaft. Bei quadrupeder Fortbewegung wurde das Körpergewicht mindestens auf je eine Vorder- und eine Hinterextremität verteilt, die Bodenkontakt hatten. Kletterbewegungen waren zwar möglich, die geringe Ausprägung der Krallen spricht jedoch dagegen, dass dies ein wesentliches Verhalten darstellte.
Bei Störung oder erhöhter Erregung konnte Eudibamus in eine schnellere Gangart übergehen und dabei Rumpf und Kopf nach hinten verlagern. Der Körper kippte dabei gleichsam wie eine Wippe aus einer horizontalen in eine stärker aufgerichtete Position. Die Hinterextremitäten und Füße waren ausreichend lang, um die veränderte Gewichtsverteilung bis zum Körperschwerpunkt zu tragen. Diese Haltung ermöglichte eine zweibeinige Fortbewegung, bei der der Vortrieb durch eine kontrollierte Vorverlagerung der Körpermasse bei jedem Schritt erzeugt wurde – vergleichbar mit heutigen zweibeinigen Organismen. Eudibamus „fiel“ dabei gewissermaßen in den jeweils nächsten Schritt hinein.
Im Gegensatz zu anderen Bromacker-„Ursauriern“ wie Orobates, Diadectes oder Dimetrodon konnten Eudibamus bislang keine eindeutig zuzuordnenden Fährten zugewiesen werden. Die Spurenfossilien des Ichnotaxons Tambachichnium stellen jedoch einen möglichen Kandidaten für Eudibamus oder sehr ähnliche Tiere dar, auch wenn deren Urheber mindestens doppelt so groß gewesen sein müssen. Dies könnte darauf hindeuten, dass es sich bei dem bekannten Holotyp von Eudibamus cursoris um ein juveniles Individuum handelt.


Stoffwechsel und Ökologie
Die Rekonstruktionsmodelle liefern neue Erkenntnisse zur Biologie und funktionellen Morphologie von Eudibamus. Das Auftreten zweibeiniger Fortbewegung vor etwa 280 Millionen Jahren wirft grundlegende Fragen nach ihrer adaptiven Bedeutung auf – etwa im Hinblick auf Feindvermeidung oder Beuteerwerb. Daraus ergibt sich die weiterführende Frage, welche Rückschlüsse sich insgesamt auf Morphologie und Lebensweise von Eudibamus ziehen lassen.
Das Prinzip der energetischen Effizienz geht davon aus, dass Organismen ihren Energieaufwand in Relation zu den metabolischen Kosten optimieren. Dieses Konzept bietet einen wichtigen Rahmen, um die Evolution der Zweibeinigkeit bei Eudibamus zu verstehen.
Eudibamus wurde bislang als Pflanzenfresser beschrieben (D. S. Berman et al., 2000), analog zu allen anderen bekannten Vertretern der Bolosauridae. Diese Einordnung stützte sich auf die charakteristische Bezahnung mit stiftförmigen Vorderzähnen und stumpf-konischen Backenzähnen, die als Anpassung an pflanzliche Nahrung interpretiert wurde. Innerhalb der Bolosauridae nimmt Eudibamus jedoch eine Sonderstellung ein, da er die einzige Art mit erhaltenem postkranialem Skelett ist. Alle anderen Funde dieser Gruppe beschränken sich auf Schädelreste; insgesamt sind Bolosauriden extrem selten fossil überliefert.
Die Untersuchung des Skeletts von Eudibamus zeigt einen auffallend kleinen und schmalen Brustkorb, der keinen Raum für den voluminösen Verdauungstrakt und die langen Därme bietet, die für die Verarbeitung energiearmer Pflanzenkost erforderlich wären. Im Unterperm bestand die Vegetation ausschließlich aus Gymnospermen wie Farnen, Bärlappgewächsen und frühen Nadelbäumen. Gräser, zuckerreiche Blüten oder energiereiche Früchte existierten nicht. Auch der Fossilbefund des Bromacker-Ökosystems weist lediglich spärliche Reste kleinwüchsiger Pionierpflanzen (z. B. Callipteris, Walchia) auf, was Zweifel an einer rein pflanzlichen Ernährungsweise von Eudibamus aufkommen lässt.
Das agile Tier hatte einen hohen Energiebedarf, während sein kleiner, schlanker Körper nur begrenzte Möglichkeiten zur Energiespeicherung in Form von Fettreserven bot. Moderne pflanzenfressende wechselwarme Wirbeltiere, etwa Iguana iguana, ergänzen ihre blattbasierte Nahrung durch zuckerreiche Früchte und nehmen regelmäßig auch Insekten oder andere energiereiche tierische Nahrung auf. Diese Arten leben jedoch in warmen Regionen mit geringen Temperaturschwankungen – ein deutlicher Gegensatz zum extremen kontinentalen Klima der Bromacker-Region mit heißen Tagen, frostigen Nächten und regelmäßigen Monsunereignissen.
Eine geeignete Nahrungsquelle für ein Tier mit der Anatomie von Eudibamus musste daher:
– energiereich und dauerhaft verfügbar sein
– klein genug, um im Ganzen geschluckt zu werden
– für größere Konkurrenten von geringem Interesse sein
Die permzeitliche Landschaft des Bromackers war geprägt von Überschwemmungsereignissen, Auenflächen mit Restgewässern und spärlicher Vegetation. Funde großer Farnwedel deuten auf nahegelegene Wälder hin. Fließgewässer transportierten zahlreiche Insekten – lebend wie tot – in sogenannte Driftlinien. Zersetzendes Pflanzenmaterial bildete eine ideale Grundlage für die Entwicklung und Massenvermehrung verschiedenster Insekten und Gliederfüßer, was durch zahlreiche Fossilfunde eindrucksvoll belegt ist.
Diese reichlich vorhandene Energiequelle diente sehr wahrscheinlich zumindest teilweise als Nahrung für Eudibamus. Seine spezialisierte Bezahnung eignete sich gut zum Zerkleinern der chitinhaltigen Körper von Käfern, Schaben und anderen Insekten. Damit besetzte Eudibamus eine besondere ökologische Nische, vergleichbar mit der heutiger Watvögel unterschiedlicher Größe. Wie diese war er beweglich, zu kurzen Sprints und möglicherweise auch zu Sprüngen fähig, um Insekten zu erbeuten. Eine solche energiereiche Ernährung hätte die hohe Mobilität ermöglicht, die für zweibeinige Fortbewegung in strukturreichen Überschwemmungslandschaften und dichter Bodenvegetation erforderlich war, und könnte zudem eine erhöhte kognitive Leistungsfähigkeit begünstigt haben – etwa zur Feindvermeidung, möglicherweise gegenüber Prädatoren wie Dimetrodon.




Hautstruktur
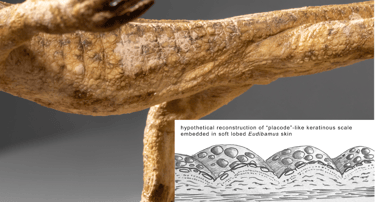
Von Eudibamus cursoris sind bislang keine fossil erhaltenen Hautreste bekannt. Wahrscheinlich besaß die Art eine spezialisierte Haut, die an die extremen, periodisch wechselnden Umweltbedingungen zwischen feucht und trocken sowie kühl und heiß angepasst war. Die seitlichen Körperpartien könnten weich und lappenartig ausgebildet gewesen sein und unter feuchten Bedingungen eine amphibienähnliche Hautatmung ermöglicht haben.
Die frei liegenden Körperoberflächen von Eudibamus vereinten vermutlich bereits weiterentwickelte Merkmale mit ursprünglich-plesiomorphen Eigenschaften. Frühe Wirbeltiere, darunter Fische und Amphibien, besaßen knöcherne Hautschuppen. Bei Eudibamus lassen sich jedoch keinerlei Hinweise auf solche Osteoderme feststellen. Keratinhaltige Schuppen entwickelten sich vermutlich im Verlauf des Perms als schützende Integumentstrukturen, die funktional den ursprünglichen knöchernen Schuppen entsprachen. Die Entstehung keratinhaltiger Schuppen – ein zentrales Merkmal der Sauropsiden – ist im Fossilbericht bislang nur unzureichend dokumentiert. Diese Lücke erschwert eine präzise Einordnung innerhalb der phylogenetischen Systematik.
Bei der Klassifikation eines Skeletts als amphibisch, reptilisch oder parareptilisch ist es entscheidend, die evolutionären Übergänge zwischen den Wirbeltiergruppen zu berücksichtigen. Merkmale moderner Reptilien, wie eine vollständig keratinisierte Schuppenhaut, entstanden nicht abrupt. Der Übergang von einer amphibienartigen zu einer reptilienartigen Haut vollzog sich sehr wahrscheinlich schrittweise über einen langen Zeitraum, mit mehreren Zwischenstufen zwischen der Haut eines Amphibiums und der eines heutigen Reptils.
Die Rekonstruktionsmodelle des Autors verfolgen das Ziel, eine solche Übergangsform sichtbar zu machen. Zur Rekonstruktion früher Keratinschuppen wurden sogenannte „Placoden“-Bildungen in Embryonen von Schildkröten und Echsen untersucht. Diese atavistischen Strukturen bestehen aus rundlichen Keratinschilden, die in die Haut eingebettet sind und sich während der Ontogenese zu typischen Schuppen weiterentwickeln. Sie dienten als Inspiration für die homogene Hauttextur, die im Modell von Eudibamus dargestellt ist. Krallen, Finger, Zehen, Unterschenkel und der Kopf waren vermutlich die am stärksten exponierten und mechanisch belasteten Körperbereiche, insbesondere durch Laufen und Rennen. Diese Zonen dürften daher am stärksten keratinisiert gewesen sein – vergleichbar mit heutigen Reptilien, auch wenn Eudibamus selbst noch keines war.
Einzelne Elemente der Lebendrekonstruktion von Eudibamus cursoris beruhen auf künstlerischer Interpretation. Sämtliche Darstellungen stehen jedoch im Einklang mit den fossil überlieferten Befunden, dem bekannten Skelettbau sowie dem rekonstruierten Lebensraum, wie er durch aktuelle Forschung belegt ist. Insgesamt ermöglicht diese Arbeit einen außergewöhnlich detaillierten Einblick in ein längst ausgestorbenes Tier – bemerkenswert angesichts des Alters von rund 280 Millionen Jahren, das die Fossilien vom Bromacker aufweisen.

Referenzen
Berman, D. S.; Reisz, R. R.; Scott, D.; Henrici, A. C.; Sumida, S. S.; Martens, T. (2000):
Early Permian bipedal reptile.
Science, 290 (5493), 969–972.
Berman, D. S.; Reisz, R. R.; Scott, D.; Henrici, A. C.; Sumida, S. S.; Martens, T. (2021):
The early Permian bolosaurid Eudibamus cursoris: earliest reptile to combine parasagittal stride and digitigrade posture during quadrupedal and bipedal locomotion.
Alibardi, L. (1996):
Scale morphogenesis during embryonic development in the lizard Anolis lineatopus.
Journal of Anatomy, 188, 713–725.
Alibardi, L. (1998):
Differentiation of the epidermis during scale formation in lizard embryos.
Journal of Anatomy, 192, 173–186.
Chang, C.; Wu, P.; Baker, R. E.; Maini, P. K.; Alibardi, L.; Chuong, C.-M. (2009):
Reptile scale paradigm: evo-devo, pattern formation and regeneration.
International Journal of Developmental Biology, 53 (5–6), 813–826.
Di-Poï, N.; Milinkovitch, M. C. (2016):
The anatomical placode in reptile scale morphogenesis indicates shared ancestry among skin appendages in amniotes.
Science Advances, 2.
weitere Artikel:
Kontaktieren Sie uns.
+49 176 61287877
brabast@gmail.com
gesponsert von: