

Another glimpse of Permian terrestrial locomotion: EUDIBAMUS CURSORIS
This article describes the scientific reconstruction of Eudibamus cursoris by Sebastian Brandt—a 280-million-year-old Permian terrestrial vertebrate from the Bromacker fossil site in Thuringia. It presents Eudibamus as the earliest known bipedal runner to date and reconstructs its anatomy, locomotion, skin structure, and ecology based on detailed fossil analyses. The work is grounded in innovative modeling techniques that consistently follow an actualistic approach aligned with current paleontological research.
Sebastian Brandt


Introduction
Eudibamus is an extinct genus of the family Bolosauridae that evolved during the Permian, approximately 280 million years ago. These enigmatic vertebrates were assigned to the Parareptilia ('reptile-like') because, despite their great age, they exhibit reptiliomorph anatomical features and thus predate known reptiles. The Parareptilia represent a unique evolutionary experiment that emerged even before the divergence of vertebrates into amphibians, reptiles, and mammals.
The only known skeleton of Eudibamus cursoris was discovered in the renowned Bromacker quarry in Thuringia (Central Germany), one of the world's most significant fossil sites for early terrestrial vertebrates.

The Bromacker finds document a complex terrestrial ecosystem of the Middle Permian with a diverse fauna and flora, including early land-dwelling vertebrates such as Orobates, Diadectes, Dimetrodon, and other species. The sandstone and mudstone sediments of the site originated on a continental high plateau characterized by periodic watercourses, ponds, and floodplains.
This Permian region was situated far from marine influences and was exposed to an extreme continental climate with strong daily temperature fluctuations. It existed during the time of the supercontinent Pangaea, long before it broke apart into today's landmasses. Evident periodic temperatures below 0 °C show that the fauna and flora developed specific anatomical adaptations to survive these climatic extremes. The vertebrate fauna of this time and region, often referred to as 'Ursaurier' (primeval saurians), does not represent direct ancestors of the later dinosaurs, but it does represent the first clearly documented, fully terrestrial vertebrates in evolutionary history.


Data basis for the reconstruction
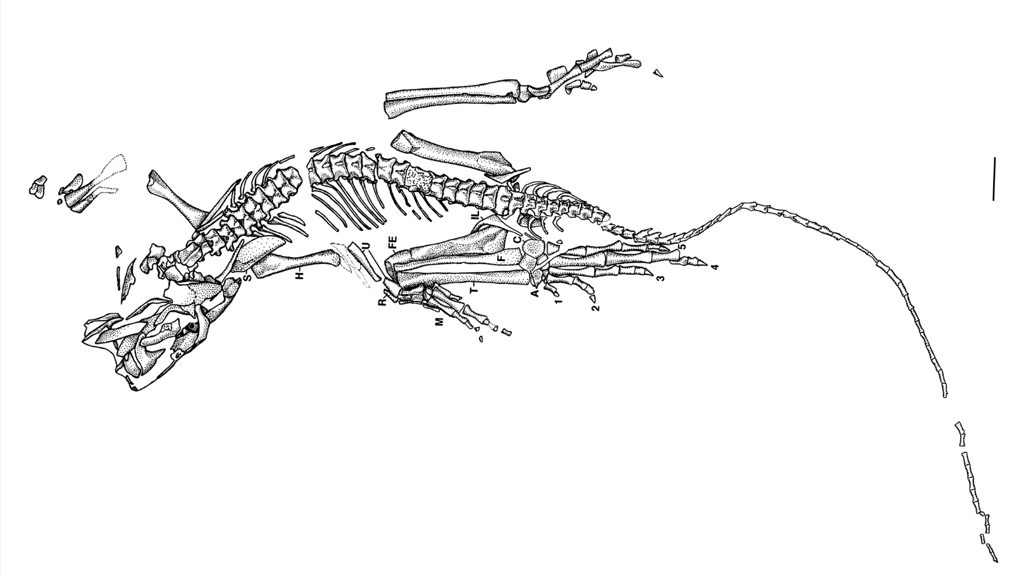
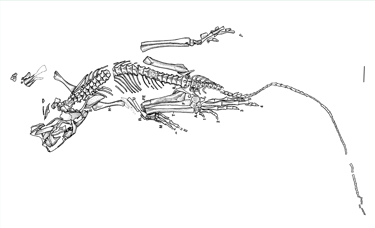
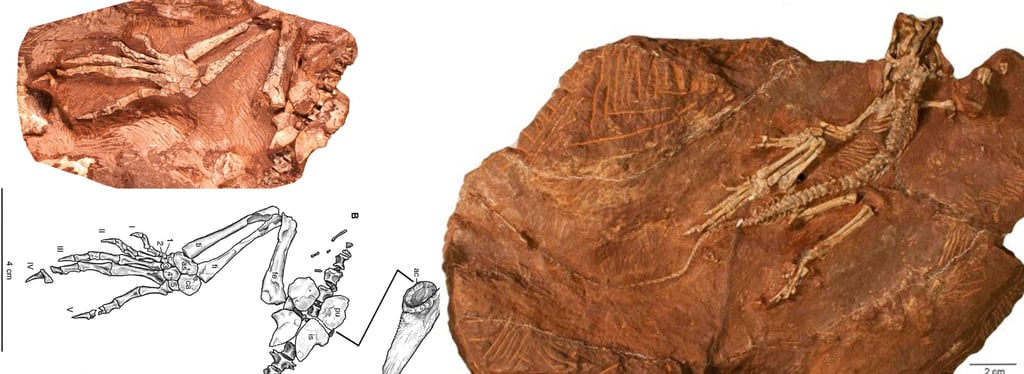
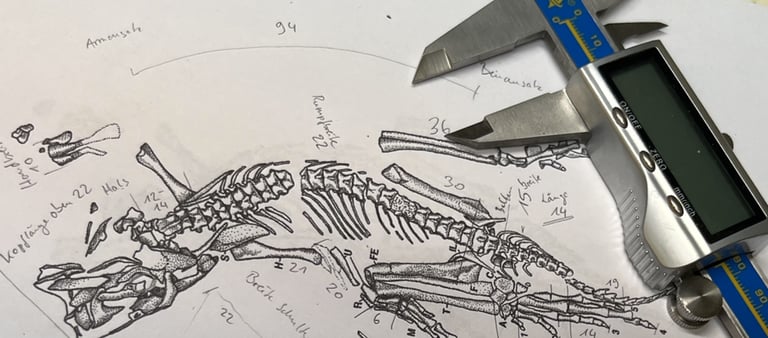
The holotype of Eudibamus cursoris (MNG 8852, Friedenstein Foundation Gotha, Germany) was discovered in 1993. It consists of an almost complete, articulated skeleton, exceptionally well preserved apart from a slightly disarticulated and diagenetically altered skull. A second specimen (MNG 12895) includes a well-preserved pelvis and an articulated hind limb. All anatomical measurements from these finds were transferred to the reconstruction at a 1:1 scale.
Key anatomical features of Eudibamus cursoris include:
– Small, slender, and highly agile body
– Long, narrow torso with short ribs and a tubular ribcage
– Slender forelimbs with elongated fingers and small claws
– Long, slender hind limbs with extended lower legs and only weakly clawed toes
– Feet with a developing heel structure
– Short, robust neck without cervical ribs
– Triangular skull with large eye sockets
– Specialized dentition with peg-like anterior teeth and conical posterior teeth
– Slender jaw bones
– Long, slender tail with strong proximal ribs
– Very small pelvis with diagonally oriented hip joints
– Clear skeletal parallels to the extant Physignathus cocincinus
Overall, the skeletal morphology indicates an extremely lightweight build, suggesting high mobility and specialized terrestrial locomotion.
The reconstruction models incorporate ecological and biological insights developed through extensive research and interdisciplinary collaboration with paleontologists including Dr. Thomas Martens, Dr. Tom Hübner, Dr. Ralf Werneburg, and other specialists.
Reconstruction of functional morphology
Locomotion
Berman et al. (2000) identified Eudibamus cursoris as the earliest known vertebrate capable of bipedal locomotion. For this study, two reconstruction models were developed: one in a quadrupedal posture, interpreted as the primary mode of movement, and a second depicting a somewhat unsteady bipedal run. The analysis draws on motion studies of extant species, in particular the unusual bipedal running behavior of Basiliscus plumifrons, to better understand the functional shift between locomotor modes.
The primary mode of locomotion in Eudibamus was quadrupedal. The animal likely moved along muddy shorelines and through early vegetation in the Bromacker landscape. During quadrupedal movement, body weight was supported by at least one forelimb and one hind limb in contact with the ground. Climbing may have been possible, but the weak development of the claws suggests it was not a dominant behavior.
Under disturbance or heightened activity, Eudibamus could shift into a faster gait by moving its body mass backward. The torso and head tilted upward, transitioning from a horizontal to a more upright position. The elongated hind limbs and feet were capable of supporting this shift in balance. This posture allowed for bipedal locomotion, with forward motion generated by a controlled forward shift of body mass with each step, similar to modern bipedal animals. In effect, Eudibamus moved by “falling” into the next step.
Unlike other Bromacker taxa such as Orobates, Diadectes, or Dimetrodon, no trackways can be definitively assigned to Eudibamus. However, trace fossils of the ichnotaxon Tambachichnium represent a possible match for Eudibamus or closely related forms, although the trackmakers appear to have been at least twice as large. This may indicate that the known holotype of Eudibamus cursoris represents a juvenile individual.


Metabolism and ecology
The reconstruction models provide new insights into the biology and functional morphology of Eudibamus. The appearance of bipedal locomotion around 280 million years ago raises fundamental questions about its adaptive value, particularly in relation to predator avoidance and foraging strategies. This leads to a broader question: what can be inferred about the overall morphology and lifestyle of Eudibamus?
The principle of energetic efficiency assumes that organisms optimize energy expenditure relative to metabolic cost. This concept offers a useful framework for understanding the evolution of bipedalism in Eudibamus.
Eudibamus has traditionally been described as herbivorous, in line with other members of the Bolosauridae. This interpretation is based on its dentition, with peg-like anterior teeth and blunt, conical posterior teeth, typically associated with plant-based diets. Within the group, however, Eudibamus is unique in preserving a postcranial skeleton. All other bolosaurid finds are limited to skull material, and the group is generally rare in the fossil record.
The skeletal anatomy of Eudibamus reveals a notably small and narrow ribcage, offering little space for the large digestive system and elongated intestines required to process low-energy plant material. During the Lower Permian, vegetation consisted primarily of gymnosperms such as ferns, lycophytes, and early conifers. Grasses, sugar-rich flowers, and energy-dense fruits did not yet exist. The fossil record of the Bromacker ecosystem likewise shows only sparse remains of small pioneer plants, raising doubts about a strictly herbivorous diet.
The animal’s agile build implies a relatively high energy demand, while its small, slender body would have limited its capacity to store energy reserves. Modern herbivorous ectothermic vertebrates, such as Iguana iguana, supplement a leaf-based diet with fruits or animal protein. These species, however, inhabit stable, warm climates, in contrast to the strongly seasonal conditions of the Bromacker region, with hot days, cold nights, and periodic monsoonal events.
A suitable food source for an animal with this anatomy would therefore need to be:
– energy-rich and consistently available
– small enough to be swallowed whole
– of little interest to larger competitors
The Permian landscape of the Bromacker was shaped by periodic flooding, floodplains with residual water bodies, and sparse vegetation. Finds of large fern fronds indicate nearby forested areas. Flowing water would have concentrated insects, both living and dead, along drift lines. Decomposing plant material provided ideal conditions for the proliferation of insects and other arthropods, as confirmed by numerous fossil finds.
This abundant resource likely formed at least part of the diet of Eudibamus. Its specialized dentition would have been well suited to processing chitinous prey such as beetles, cockroaches, and other insects. Eudibamus may therefore have occupied a distinct ecological niche, comparable to that of modern wading birds. Mobility, short bursts of speed, and possibly jumping would have allowed it to capture prey efficiently. Such an energy-rich diet would support the high level of activity required for bipedal movement in structurally complex floodplain environments, and may also have supported enhanced behavioral flexibility, including predator avoidance in the presence of large predators such as Dimetrodon.




Skin structure
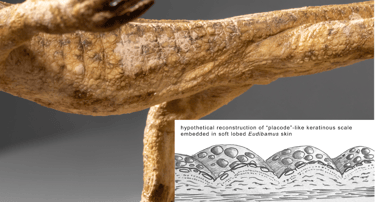
No fossilized skin remains of Eudibamus cursoris are known to date. The species likely possessed a specialized integument adapted to strongly fluctuating environmental conditions, ranging between wet and dry, and cool and hot. The lateral body surfaces may have been soft and somewhat lobe-like, potentially allowing amphibian-like cutaneous respiration under humid conditions.
The exposed body surfaces of Eudibamus likely combined more derived features with primitive, plesiomorphic traits. Early vertebrates, including fishes and amphibians, possessed bony dermal scales, yet no evidence of such osteoderms has been identified in Eudibamus. Keratin-based scales probably evolved during the Permian as protective integumentary structures, functionally replacing earlier bony scales. The emergence of keratinized scales, a key feature of sauropsids, remains poorly documented in the fossil record, complicating precise phylogenetic interpretation.
Any classification as amphibian, reptile, or parareptile must take these evolutionary transitions into account. Characteristics of modern reptiles, such as a fully keratinized scaly skin, did not appear abruptly. The transition from amphibian-like to reptilian skin was almost certainly gradual, involving multiple intermediate stages.
The reconstruction aims to make such a transitional condition visible. For this purpose, early keratinous structures were studied in the form of so-called “placodes” in embryos of turtles and lizards. These atavistic features consist of rounded keratinous elements embedded in the skin, which later develop into typical scales. They served as a model for the homogeneous skin texture shown in the reconstruction of Eudibamus. Areas subject to greater mechanical stress—such as claws, digits, lower limbs, and the head—were likely more strongly keratinized, similar to modern reptiles, although Eudibamus itself had not yet reached that stage.
Certain aspects of the life reconstruction involve informed interpretation. All elements remain consistent with the fossil evidence, known skeletal morphology, and the reconstructed habitat as supported by current research. The result offers an unusually detailed insight into an organism that lived around 280 million years ago, as documented by the Bromacker fossils.

References
Berman, D. S.; Reisz, R. R.; Scott, D.; Henrici, A. C.; Sumida, S. S.; Martens, T. (2000):
Early Permian bipedal reptile.
Science, 290 (5493), 969–972.
Berman, D. S.; Reisz, R. R.; Scott, D.; Henrici, A. C.; Sumida, S. S.; Martens, T. (2021):
The early Permian bolosaurid Eudibamus cursoris: earliest reptile to combine parasagittal stride and digitigrade posture during quadrupedal and bipedal locomotion.
Alibardi, L. (1996):
Scale morphogenesis during embryonic development in the lizard Anolis lineatopus.
Journal of Anatomy, 188, 713–725.
Alibardi, L. (1998):
Differentiation of the epidermis during scale formation in lizard embryos.
Journal of Anatomy, 192, 173–186.
Chang, C.; Wu, P.; Baker, R. E.; Maini, P. K.; Alibardi, L.; Chuong, C.-M. (2009):
Reptile scale paradigm: evo-devo, pattern formation and regeneration.
International Journal of Developmental Biology, 53 (5–6), 813–826.
Di-Poï, N.; Milinkovitch, M. C. (2016):
The anatomical placode in reptile scale morphogenesis indicates shared ancestry among skin appendages in amniotes.
Science Advances, 2.