

Sturia brandti n. sp. and Sephardonautilus nov. gen.
This paper examines the Upper Muschelkalk as a newly emerged epicontinental sea ecosystem of the Middle Triassic, demonstrating how ammonoid immigrations from Alpine and Sephardic faunal provinces enable precise biostratigraphic and chronostratigraphic correlations. Based on newly described taxa—Sturia brandti n. sp. (discovered by Sebastian Brandt) and Sephardonautilus nov. gen.—migration routes, allopatric speciation processes, and evolutionary relationships are reconstructed. The findings provide a more refined temporal subdivision of the Upper Muschelkalk and offer new insights into faunal dynamics during the Anisian/Ladinian transition.
Sigfried Rein, Sebastian Brandt


Sturia brandti n. sp. and Sephardonautilus nov. gen. – Immigrants, Migration Routes, and Correlations in the Upper Muschelkalk (Middle Triassic)
Abstract
As a newly emerged ecosystem, the Upper Muschelkalk represents a tectonically dominated epicontinental sea. Immigrated macrofossils from Alpine and Sephardic faunal provinces enable biostratigraphic and chronostratigraphic correlation. During the lower trinodosus zone, Germanonautilus salinarius migrated from the Austroalpine-Dinaric faunal province into the newly formed inland sea via a Helvetic-Alemannic gateway. Germanonautilus bidorsatus emerged cladogenetically from this ancestral species. Following the same route during the middle trinodosus zone, Sturia sansovinii migrated from the Austroalpine faunal province and cladogenetically formed the new species Sturia brandti n. sp.
The most significant step in the colonization of the Upper Muschelkalk ecosystem occurred during the lower Illyrian from the Sephardic faunal province through a uniquely and briefly tectonically opened Burgundian-Rhenish gateway. With Sephardonautilus tridorsatus nov. comb. as the type species of the new genus Sephardonautilus nov. gen. and the ancestral species Sephardonautilus salinarius, the phylogenetic link between the individuals of the Muschelkalk sea and the Sephardic faunal province is documented. The same applies to the evolutionary species Ceratites nodosus, which emerged after allopatric speciation from an unknown species of the genus Ceratites (de Haan) of the Sephardic faunal province.
The ancestral species Sephardonautilus salinarius from the trinodosus zone of the upper Gevanim Formation of Makhtesh Ramon chronologically confirms the correlation of the migration route of Sephardonautilus tridorsatus. The morphocline of the unprecedentedly eurypotent evolutionary species Ceratites nodosus allows for a seamless biostratigraphic subdivision of the Upper Muschelkalk.
Populations of the Southern Alpine morphs Flexoptychites angusto-umbilicatus and Protrachyceras recubariense hypothetically immigrated concurrently with the brachiopod Punctospirella fragilis. The timing of this fourth immigration phase corresponded chronostratigraphically and biostratigraphically to the Anisian/Ladinian transition. Through physiological adaptation, Flexoptychites and Protrachyceras survived in SW Germany from the evolutus zone into the spinosus zone. They await a new cladogenetic species status.
For the definition of the Anisian/Ladinian boundary in the Upper Muschelkalk, the Spiriferina bed would mark the transition from the compressus zone to the evolutus zone. This correlates with the transition from the secadensis zone to the curionii zone in the Southern Alpine faunal province.
Immigrants from the Ladinian of the Schreyeralm Formation, Parapinacoceras thiemei and Gymnites bruneli, briefly colonized the basin interior in Thuringia. Through the allopatric speciation of an unknown species of the genus Ceratites (de Haan) from the Sephardic faunal province, the sister species Ceratites nodosus and Ceratites tornquisti emerged. The phylogenesis of the species Ceratites tornquisti began in the lower Illyrian in the Palaeotethys and ended in the Longobardian within the Thuringian Grenzdolomit as the chronospecies Ceratites schmidti. The clarification of the previously unknown phylogenesis of Ceratites tornquisti in the habitats of Dobruja, Sardinia, Vicentin, Provence, and Thuringia remains reserved for detailed analysis.
1. Introduction
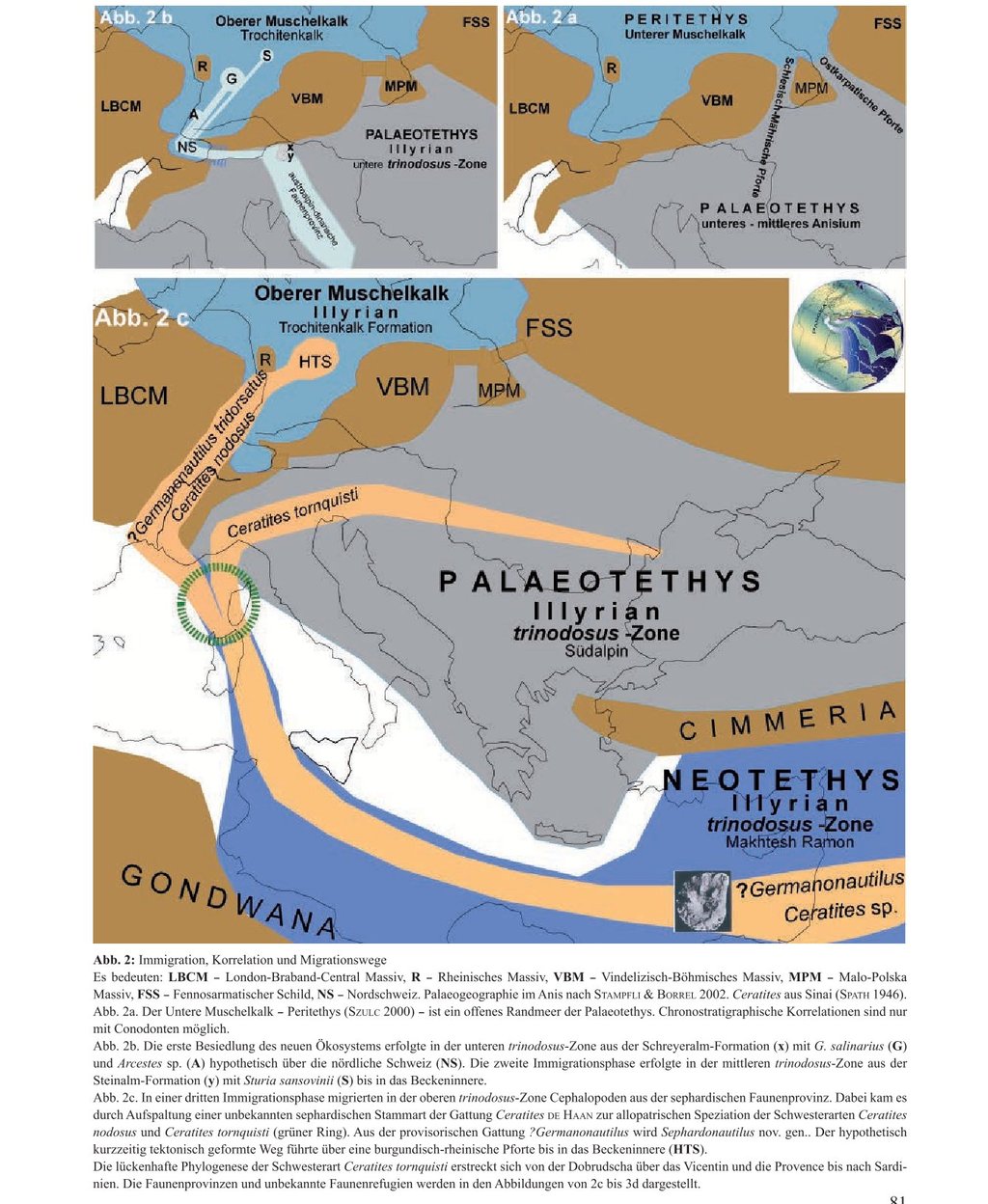
During the Middle Triassic, present-day Central Europe was a depression separated from the Palaeotethys Ocean to the southeast by the Bohemian-Vindelician Highland and the Carpathian Foreland. Driven by epeirogenic and eustatic factors, it was flooded first through the East Carpathian gateway and later through the Silesian-Moravian gateway (Fig. 2a). Thus, the Peritethys emerged as a shallow marginal sea of the Palaeotethys during the Lower Muschelkalk (Szulc 2000). With a closed northern coastline, it extended from Poland across Germany to France and the northern border of Switzerland. Until the middle Anisian, faunal migration from the Asian and Alpine faunal provinces of the Palaeotethys proceeded unhindered through these gateways (Kozur 1974). The palaeogeographic position of these migration routes from the directly adjacent Palaeotethys at the southeastern margin into this marginal sea is relatively well determined.
However, the fossil-poor carbonates of the Lower Muschelkalk do not permit a basin-wide biostratigraphy using macrofossils. Consequently, due to the lack of biostratigraphic reference profiles, chronostratigraphic correlations with the Tethys region are only possible using conodonts (Narkiewicz & Szulc 2004).
Following the closure of the eastern gateways, the marine connection to the Palaeotethys was completely severed. Due to the extreme increase in salinity caused by high evaporation rates, hospitable conditions largely vanished during the Middle Muschelkalk for a period of over 1.1 million years.
2. The Upper Muschelkalk
In contrast to the paleogeographically well-defined eastern gateways of the marginal sea during the Lower Muschelkalk, all interpretations of gateway openings to the Tethys Ocean during the Upper Muschelkalk remains hypothetical. However, unlike the Lower Muschelkalk, the carbonates of the Upper Muschelkalk are unusually fossiliferous. Using macrofossils, they ensure the establishment of a high-resolution biostratigraphy and enable a chronological documentation of the newly emerged ecosystem within the Germanic faunal province. Following their immigration, the evolutionary development of the organisms occurred under endemic conditions. The immigration of a species ended with reproductive isolation from its ancestral species and the cladogenetic emergence of a new species.
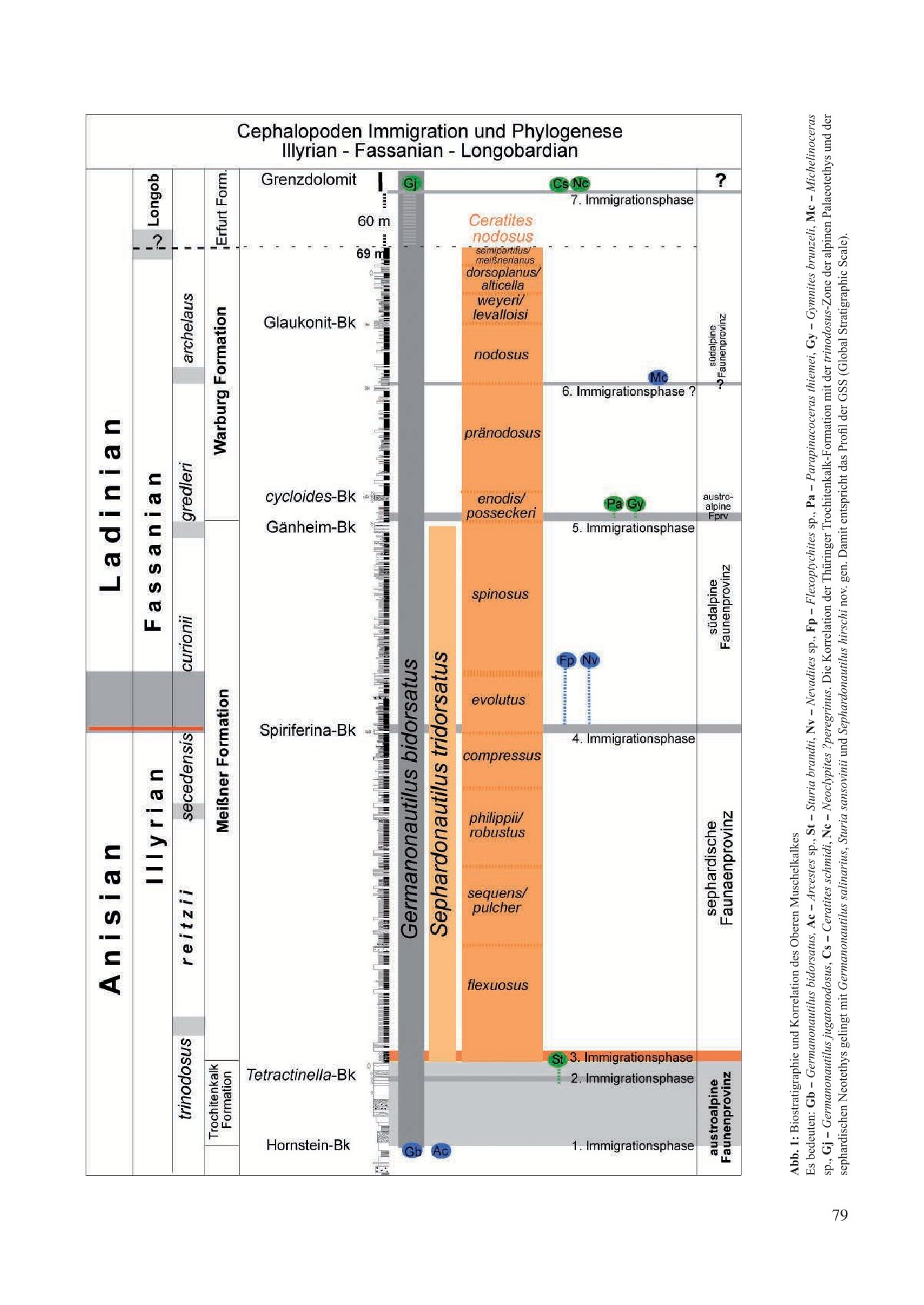
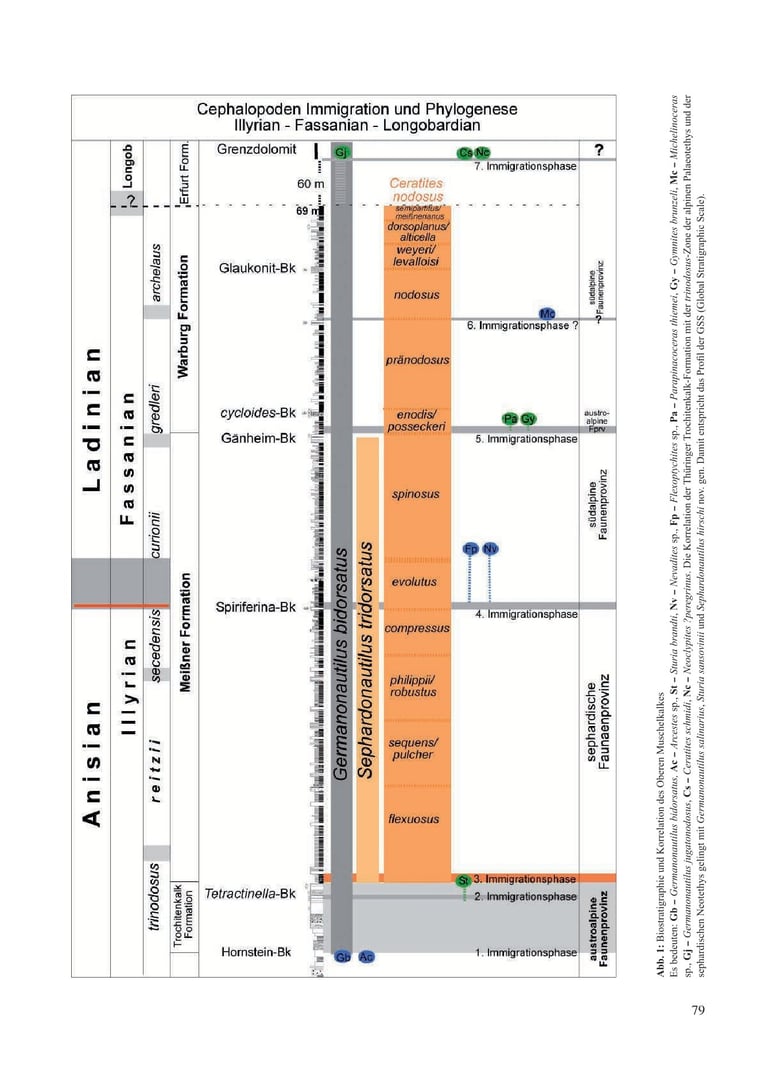
The biostratigraphic profile, supported by macrofossils, documents seven immigration phases into this inland sea, leading to new speciations (Fig. 1). As a chronological record, these phases enable correlation with various Tethyan faunal provinces and the reconstruction of hypothetical migration routes of the immigrants.
In northern Switzerland, the Upper Muschelkalk is fully developed as the Schinznach Formation (Pietsch et al. 2016; Adams et al. 2019). Macrofossils document the southwesternmost extent of the Muschelkalk sea (Merki 1961). In the far east of Switzerland lies the St. Gallen Rhine Valley, bordering Austria and Liechtenstein. A connection via the Alpine Rhine depression to the western margin of the Palaeotethys therefore appears geologically plausible ("Western Gate," Szulc 2000). This would represent the shortest migration route along the eastern edge of the Black Forest through southwestern Germany into the basin ("Alemannic Gateway," Bachmann et al. 2010).
Hitherto, a marine connection to the south via a "Burgundian Gateway" has been favored. At the very least, the cephalopods that immigrated from the Sephardic faunal province of the Neotethys into the Muschelkalk sea must have used this Burgundian route between the Vosges and the western edge of the Black Forest via a "Rhenish Gateway" (Rein 2016).
3. The Immigration Phases
3.1 First Immigration Phase (Fig. 1, Fig. 2b/x)
Middle Muschelkalk, Diemel Formation
Rhyncholithes hirundo (Germanonautilus) (Gb): Location Oßmaritz near Jena, Thuringia, underlying the Upper Hornsteinbank (Knaust 1991).
Arcestes sp. (Ac): Location Weil der Stadt, eastern edge of the Black Forest, Southwest Germany, underlying the Hornsteinbank (Hohenstein 1913; Schmidt 1928).
3.2 Second Immigration Phase (Fig. 1, Fig. 2b/y)
Upper Muschelkalk, Trochitenkalk Formation, Tetractinella bed, Thuringia
Tetractinella trigonella: ecostratigraphic marker bed.
Sturia sansovinii (St): Location Geilsdorf near Stadtilm, overlying the Tetractinella bed, Thuringia.
3.3 Third Immigration Phase (Fig. 1)
Upper Muschelkalk, Meißner Formation, flexuosus zone, Thuringia
Ceratites nodosus: Hessian-Thuringian Depression (Seidel 2013), Thuringia, Troistedt, approx. 1 m above the Tetractinella bed (Rein 2007, 2017).
?Germanonautilus tridorsatus: Hessian-Thuringian Depression, Thuringia, Troistedt, approx. 1 m above the Tetractinella bed (Rein 2016).
3.4 Fourth Immigration Phase (Fig. 1)
Upper Muschelkalk, Meißner Formation, Spiriferina bed, Southwest Germany
Punctospirella fragilis: ecostratigraphic marker bed.
Flexoptychites angusto-umbilicatus (N): Location Gundelsheim, spinosus zone (Urlichs & Kurzweil 1997).
Protrachyceras recubariense (P): Location Winterrain quarry near Weissach, Southwest Germany, middle spinosus zone (Urlichs 1978).
3.5 Fifth Immigration Phase (Fig. 1)
Upper Muschelkalk, Warburg Formation, Thuringia
Pinacoceras sp. (Pa): Location Ettenhausen near Eisenach, Thuringia, Warburg Formation, enodis/posseckeri zone (Rein 2009; Rein & Werneburg 2010).
Gymnites sp. (Gy): Location Ohrdruf near Gotha, Thuringia, Warburg Formation, enodis/posseckeri zone (Rein & Werneburg 2010).
3.6 Sixth Immigration Phase (Fig. 1)
Upper Muschelkalk, Warburg Formation, SW Germany
Michelinoceras campanile (Mc): Location Trigonodus-Dolomit of Buigen near Mönsheim, SW Germany; based on the associated fauna, it likely originates from the nodosus zone (Schröder 1977; Urlichs & Schröder 1980).
3.7 Seventh Immigration Phase (Fig. 1)
Lower Keuper, Erfurt Formation, Grenzdolomit, Thuringia
Ceratites schmidti (Cs): Locations Sülzenbrücken and Reisdorf, Thuringia, Grenzdolomit (Zimmermann 1883; Philippi 1901; Müller 1969, 1970, 1973).
Neoclypites? peregrinus (Nc): Location Reisdorf, Thuringia, Grenzdolomit (Müller 1973).Germanonautilus jugatonodosus (Jn): Branchewinda, Thuringia, Grenzdolomit (Zimmermann 1889; Rein 2014, 2015; Urlichs & Mundlos 1984; Urlichs 2000, 2015).
Description
4.1 Immigration of Germanonautilus salinarius from the Austroalpine-Dinaric Faunal Province (Schreyeralm Formation) (Fig. 2b/x)
During the Middle Triassic, the Austroalpine-Dinaric faunal province, characterized by a shared fauna, extended from Han Bulog to the Schreyeralm limestones of the Salzburg region. The Schreyeralp is the type locality of Nautilus salinarius Mojsisovics, 1882 and Arcestes sp. It documents an Illyrian age via Paraceratites trinodosus. Nautilus salinarius is also described from the Schiechlinghöhe (Diener 1900) and Han Bulog (Hauer 1887).
The first immigration phase into the newly emerging habitat of the Upper Muschelkalk occurred at the base of the Illyrian within the lower trinodosus zone. The immigrated Nautilus salinarius and Arcestes sp. from the Austroalpine-Dinaric faunal province enable the first chronostratigraphic and biostratigraphic correlation with the Palaeotethys. Descriptively, the ancestral species Nautilus salinarius Mojsisovics, 1882 gave rise to the new species Nautilus bidorsatus Schlotheim, 1820. Both species were united by Mojsisovics (1902) within the newly defined genus Germanonautilus, with Germanonautilus bidorsatus designated as the type species.
Palaeogeographically, the chosen migration route via the Alpine Rhine depression remains hypothetical. However, the development of the Upper Muschelkalk in the Schinznach Formation of northern Switzerland (Pietsch et al. 2016; Adams et al. 2019) confirms the possibility of a marine connection via the Alemannic Gateway (Bachmann et al. 2010) into SW Germany.
The discovery of Rhyncholithes hirundo from the Middle Muschelkalk of Jena (Knaust 1991) documents the immigration of Germanonautilus salinarius from the Austroalpine region of the Palaeotethys during the trinodosus zone. Since the jaw is part of the soft body, post-mortem transport can be ruled out. Thus, its occurrence in the basin interior proves the successful colonization of the new habitat within the upper Diemel Formation. Furthermore, it serves as physiological evidence of the unusual eurypotent adaptability of the nautilid. The species Germanonautilus bidorsatus, which emerged following reproductive isolation, remains the only Alpine cephalopod species able to permanently integrate into the Upper Muschelkalk.
Stratigraphically, the Upper Muschelkalk in SW Germany and Thuringia begins with the Hornsteinbank. Previously, the find of Arcestes sp. from the Middle Muschelkalk of SW Germany was interpreted as a post-mortem, passively transported shell. However, the example of Germanonautilus demonstrates that active colonization of the new habitat already began in the Diemel Formation with the first influx of freshwater from the transgressing Palaeotethys. Correlated with the immigration of Germanonautilus salinarius, this corresponds to the lower trinodosus zone.
4.2 Immigration of Tetractinella trigonella and Sturia sansovinii from the Austroalpine Faunal Province (Steinalm Formation) (Fig. 2b/y)
The circumstances surrounding the immigration of the brachiopod Tetractinella trigonella and the cephalopod Sturia sansovinii differ from those of Germanonautilus salinarius. Tetractinella trigonella has not been recorded from the facies of the Schreyeralm Formation (Schreyeralp, Schiechlinghöhe) (Bittner 1890). It is known from the binodosus zone of the Eastern Alps, the Balaton Highlands, and Recoaro (Bittner 1890; Torti & Angiolini 1997; Palfy 2003). Consequently, the record of Tetractinella trigonella from the trinodosus zone of the Steinalm Formation appears significant (Broili 1927).
In contrast, Sturia sansovinii is an important index fossil of the trinodosus zone due to its wide horizontal distribution (Diener 1900). It is documented from the type locality Schreyeralmkalk (Mojsisovics 1882), the Schiechlinghöhe (Diener 1900), and the Steinalm Formation (Broili 1927; Schnetzer 1934).
A further example of its wide horizontal distribution as an index fossil of the trinodosus zone is the record of Sturia sansovinii in the Thuringian Muschelkalk. The specimen was recovered on June 24, 2016, during the construction of the B 90 federal highway near Geilsdorf (R 44 35 590; H 56 22 800). This find confirms the correlation of the upper Trochitenkalk Formation in Thuringia with the Alpine middle trinodosus zone. It therefore holds particular value for the chronostratigraphic correlation between the Germanic and Alpine Middle Triassic.
The Geilsdorf locality is situated in the basin center; stratigraphically, the find is located approximately 3.5 m above the immigration level of Tetractinella trigonella. It is highly probable that Sturia sansovinii was not an isolated individual but part of an immigrant population. Due to territorial and reproductive isolation from the Alpine ancestral species, a new cladogenetic species emerged under endemic conditions.
Systematics
Family Ptychitidae Mojsisovics, 1882
Genus Sturia Mojsisovics, 1882
Type species Sturia sansovinii Mojsisovics, 1882
Species Sturia brandti n. sp.
Description of Sturia brandti n. sp.
Holotype: Single find from 24.06.2016, locality Geilsdorf (R 44 35 590; H 56 22 800), Upper Muschelkalk, Trochitenkalk Formation, Thuringia, collection of the Natural History Museum Erfurt, collection no. NME C16;200.
Preservation: The specimen is a partially preserved internal mold (steinkern) representing approximately 120° of the phragmocone whorl. Apart from limonitic coatings on the steinkern surface as diagenetically altered remains of the hypostracum, no shell remains are fossilized. Consequently, surface structures of the lateral side are absent.
Derivatio nominis: The species name Sturia brandti n. sp. not only refers to the discoverer of the fossil but also honors the versatile achievements of Sebastian Brandt from Kornhochheim, whose meritorious work is dedicated to the Thuringian Muschelkalk.
Comparison with the original of Sturia sansovinii Mojsisovics, 1882
Size: The marginal width of the specimen is 124 mm. The whorl height increases from approx. 65 mm to 86 mm.
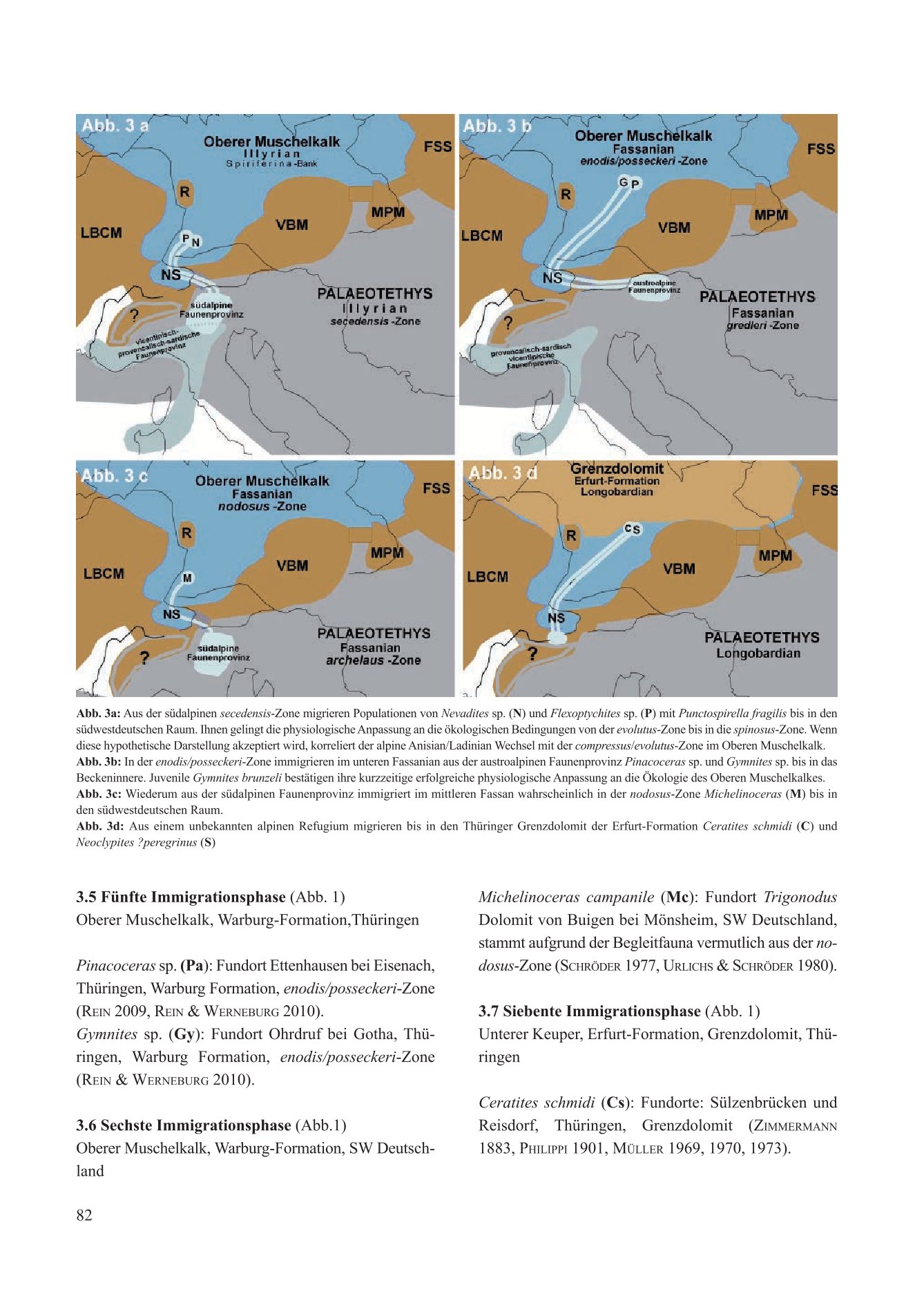
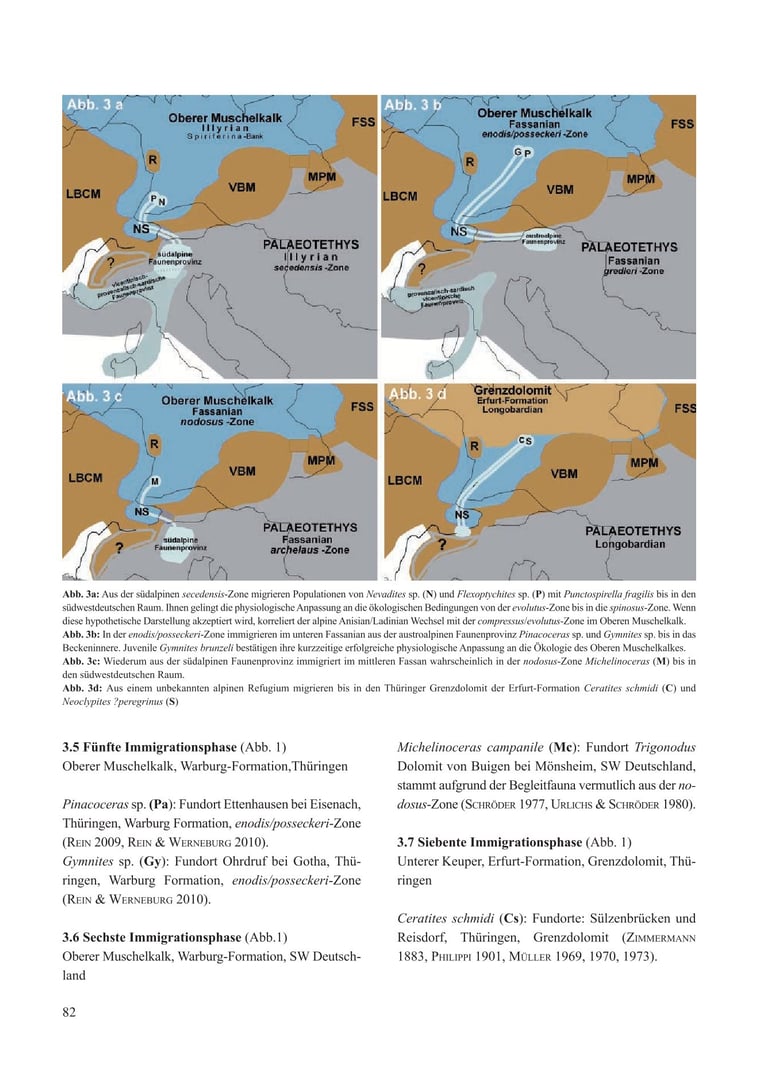
Fig. 4: Sturia brandti n. sp., Geilsdorf, lower Meißner Formation Thuringia, Germany


The whorl cross-section Q = (b/h)*100 can only be calculated approximately. It is consistently 46. The values for the whorl height of the original are 68 mm, with a shell cross-section of 41.
Mojsisovics (1882) describes the phragmocone diameter of the largest specimen available to him as 135 mm. With a reconstructed total diameter of approx. 225 mm (Fig. 6), the size of the Thuringian find is slightly larger than the Schreyeralp morphs of Mojsisovics.
Suture comparison (Fig. 5) Fig. 5
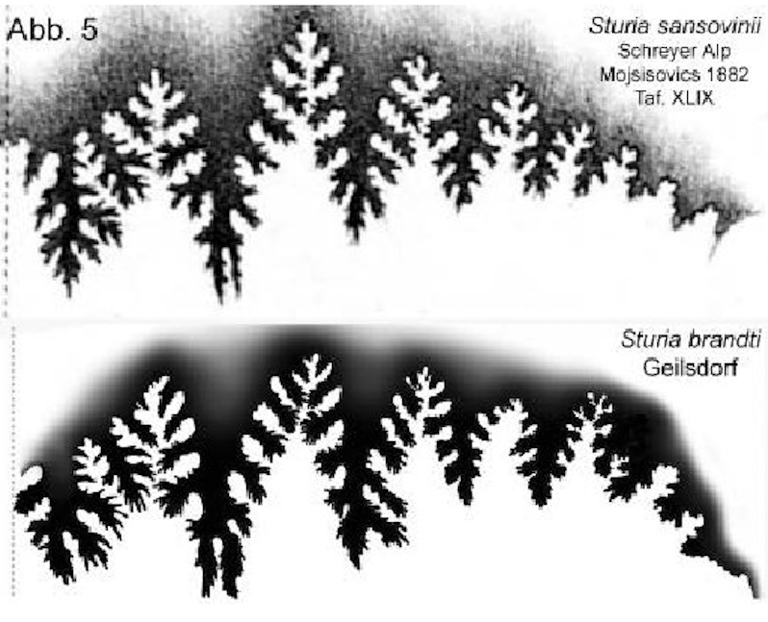
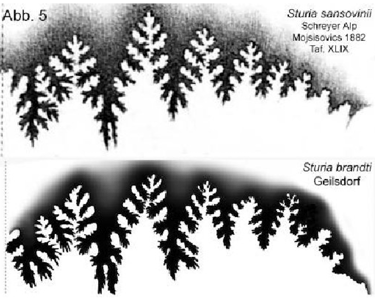


Fig. 6: Reconstruction of the shell size of Sturia brandti n. sp.
Fig. 5: Suture comparison of Sturia sansovinii Mojsisovics and Sturia brandti n. sp. [1]
Lobes: Only five auxiliary lobes are developed. The external lobe is divided by a broad median saddle that extends onto the lateral flanks. The external lobe is wider than in the original. Similarly, the first and second lateral lobes bifurcate into two tips significantly wider than in the original.
Saddles: The external saddle appears broad, as in the original, and possesses a distinct outer branch. The saddles are narrow, high, and pointed; in the upper half, they are incised down to the narrow saddle stem, while the lower part features a broad, unincised saddle stem. As in the original, the external saddle is slightly lower than the first lateral saddle.
The suture of the Thuringian specimen differs significantly from the original of Sturia sansovinii Mojsisovics, 1882, regarding the development of the lateral lobes and the saddle stems.
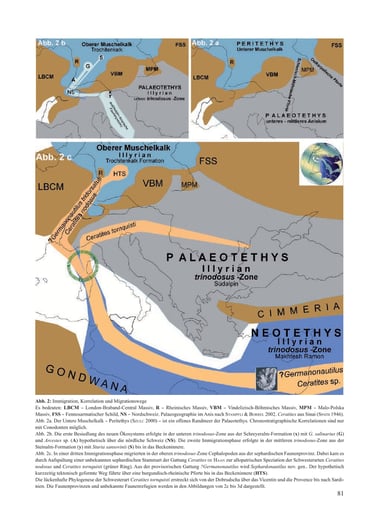
4.3. The Immigration of Sephardic Cephalopods (Fig. 2c)
The cephalopods of the third immigration phase in the basal Illyrian, ?Germanonautilus tridorsatus Böttcher, 1938 and Ceratites nodosus Schlotheim, 1820, were not species from the Alpine Tethys region but foreign forms from the Neotethys. The fossil record documents the actual timing of their immigration into the Muschelkalk sea (REIN 2008, 2016, 2017). In the Upper Muschelkalk of Thuringia, the Trochitenkalk Formation (basal Illyrian) extends from the upper Hornstein layer to one meter above the Tetractinella bed. With the facies change above this ecostratigraphic marker bed (HAGDORN & SIMON 1993), the Meißner Formation begins in Thuringia, biostratigraphically marked by the flexuosus zone with the first appearance of Ceratites. The first joint fossil record of the cephalopods ?Germanonautilus tridorsatus and Ceratites nodosus occurs within the clay/marl layers of this biozone.
This evidence confirms their joint immigration via the Rhenish Gateway (REIN 2017) into the Hessian-Thuringian Depression (SEIDEL 2013), and thus directly into the center of the Muschelkalk sea. The migration route into the Germanic faunal province is hypothetical (Fig. 2c). However, it can be traced biostratigraphically and chronostratigraphically through the genealogical correlation of ?Germanonautilus tridorsatus (REIN 2016). A statistical-biometric analysis revealed that the form ?Germanonautilus tridorsatus Böttcher, 1938—originally assigned to the genus Germanonautilus Mojsisovics, 1902—should be placed in its own genus based on morphological and physiological grounds (REIN 2016). Since the origin of the ancestral lineage of the species ?Germanonautilus tridorsatus Böttcher, 1938 lies in the Sephardic faunal province, the generic term ?Germanonautilus was initially chosen.
Kummel (1960) assigned the nautilid morphs from Areif en Naqa and Makhtesh Ramon (Sinai/Negev) with steep umbilical walls to the genus Germanonautilus Mojsisovics, 1902. Subsequently, PARNES (1986) described the nautilids from the upper Gevanim Formation of Makhtesh Ramon synonymously as Germanonautilus salinarius Mojsisovics, 1902, and likewise assigned all other Sephardic nautilid forms with steep umbilical walls to Germanonautilus. This is a misdiagnosis, as the umbilical wall of Germanonautilus slopes obliquely from the umbilical margin to the suture.
The generic term Germanonautilus Mojsisovics, 1902 is therefore invalid for the Sephardic morphs. The same applies to all other Sephardic species assigned to Germanonautilus by PARNES (1986).
The steep umbilical slope of the internal molds of Sephardic nautilid morphs is also a characteristic of the Muschelkalk genus ?Germanonautilus. Based on this shared feature of internal mold preservation, the new genus Sephardonautilus nov. gen. is established.
Due to the shared generic characteristics described above, the species ?Germanonautilus tridorsatus Böttcher, 1938 is transferred to the newly established genus Sephardonautilus nov. gen., with Sephardonautilus tridorsatus (Böttcher, 1938) designated as the type species.
Description
The whorl cross-section is highly variable, ranging from square to trapezoidal. The flanks can be smooth or shallowly convex; the slightly rounded marginal edge is smooth or, rarely, sculpted with faint bulges or nodes towards the lateral side. The ventral side is flat and may transition into a narrow, shallow groove parallel to the marginal edge. The umbilical width is variable. The umbilical margin can be developed into a strong bulge. From the umbilical margin, the umbilical wall falls steeply towards the suture. Starting from the last septum, a hyponomic sinus with a broadly rounded base begins at 115° on the ventral side of the shell. From there, the funnel margins run parallel to the marginal edge up to the ventral shell aperture at 165°. The shell aperture curves over the marginal edge in a tightly rounded arc, initially convex on the lateral side towards the rear, before switching shortly after into a wide concave arc towards the front. Following this concave arc, periodic bulging shell thickenings occur laterally. Measured through the umbilicus, the shell aperture ends at 235° in a partially strongly developed umbilical bulge and then turns perpendicular to the suture. Thus, the opening of the ventral shell aperture for the soft body extends from 115° to 235° over an angular measure of 120°.
The species Sephardonautilus tridorsatus (Böttcher, 1938) nov. comb. emerged in the lower Illyrian of the Upper Muschelkalk following reproductive isolation from the ancestral species Sephardonautilus salinarius (Parnes, 1986) and became extinct in the Lower Fassanian above the Gänheim Bed.
Systematics
Family Tainoceratidae Hyatt, 1883
Genus Sephardonautilus nov. gen.
Type species Sephardonautilus tridorsatus (Böttcher, 1938)
All taxa described by PARNES (1986) as Germanonautilus are transferred to the new genus Sephardonautilus nov. gen.:
Sephardonautilus salinarius (Parnes, 1986) nov. comb.
Sephardonautilus saharonicus (Parnes, 1986) nov. comb.
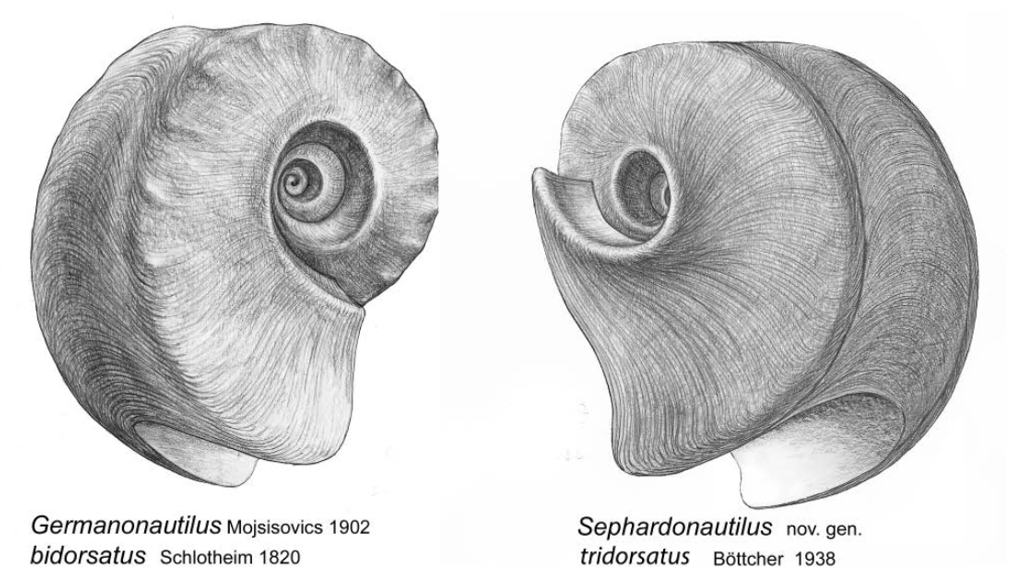
Abb. 7: Germanonautilus bidorsatus und Sephardonautilus tridorsatus
Sephardonautilus bidorsatus bidorsatus (Parnes, 1986) nov. comb.
Sephardonautilus bidorsatus jugosus (Parnes, 1986) nov. comb.
Sephardonautilus bartovi (Parnes, 1986) nov. comb.
Sephardonautilus bartovi bartovi (Parnes, 1986) nov. comb.
Sephardonautilus bartovi timidus (Parnes, 1986) nov. comb.
Sephardonautilus bartovi gravidus (Parnes, 1986) nov. comb.
Sephardonautilus ellipticus (Parnes, 1986) nov. comb.
The morphological differences between the genera Germanonautilus and Sephardonautilus nov. gen. become evident upon comparison. The new genus Sephardonautilus is defined by the expanded description of the type species Sephardonautilus tridorsatus (Böttcher, 1938) (Rein 2016). All morphological and physiological details presented in this description of the type species are due to the exceptionally good fossil preservation of the Thuringian Muschelkalk.
Ceratites sp. of the genus Ceratites de Haan, 1825
According to Parnes (1986), the uppermost part of the Gevanim Formation represents the base of the trinodosus zone and thus the lower Illyrian. Consequently, the stratigraphic level of Sephardonautilus salinarius corresponds chronostratigraphically to the third immigration phase into the Muschelkalk sea (Fig. 1c). Unfortunately, ammonites are absent in this horizon; thus, there is no fossil evidence from Israel corresponding to the ceratitid immigrants. Information regarding the morphology and genealogy of the unknown ancestral form is provided by two moderately preserved phragmocones with strong dichotomous sculpture and ceratitic structure from Aref en Naqa in the Egyptian Sinai. These were tentatively assigned to Progonoceratites as flexuose forms by Spath (1946). The evolution of the unknown species of the genus Ceratites de Haan ends with the cladogenetic split of the population at the end of its tectonically influenced joint migration route (green ring, Fig. 1c). Through this allopatric speciation, the two sister species Ceratites nodosus and Ceratites tornquisti emerged. Such paleogeographic and chronostratigraphic evidence of a speciation event serves as an evolutionary model case for the biological and evolutionary species concept in paleontology (Willmann 1985, Reif 2009). The morphocline of the endemic evolutionary species Ceratites nodosus, documented without fossil gaps, begins in the Hessian-Thuringian Depression (HTS) of the Germanic faunal province (Fig. 2c). The phylogenesis of the species Ceratites tornquisti of the genus Ceratites de Haan begins in the trinodosus zone without fossil evidence in a hypothetical Vicentine-Romanian faunal province between the Southern Alps (Recoaro) and Dobruja (Zibil), Fig. 2c. The externally imposed separation of populations resulted in reproductive isolation of individuals between the endemic inland sea (Upper Muschelkalk) and the unpredictably open Palaeotethys. The interruption of gene flow led to an uneven allele distribution of the separated populations and resulted in character divergence.
4.4. The Anisian/Ladinian Transition and Immigration from the Southern Alpine Faunal Province (Fig. 3a)
The fourth immigration phase in the upper Illyrian corresponds to the migration of the brachiopod Punctospirella fragilis and presumably the bivalve Chlamys (Praechlamys) reticulata. In the Southern Alpine faunal province, Punctospirella fragilis is recorded sporadically in the upper Anisian without recognizable index value (Bittner 1890, Torti & Angiolini 1997). In the Upper Muschelkalk, the immigrated brachiopod forms an important ecostratigraphic marker bed with the Spiriferina bed (Hagdorn & Simon 1993). It documents a real immigration event at the base of the evolutus zone. However, the finds of the two Alpine ammonites Flexoptychites angusto-umbilicatus and Protrachyceras recubariense originate from the spinosus zone of SW Germany. Since the Upper Muschelkalk was not a marginal sea open to the Palaeotethys, this renewed immigration raises several questions:
– Does the fossil horizon in the Muschelkalk really correspond chronostratigraphically and biostratigraphically to the Southern Alpine horizon of the immigrants?
– Are these passively post-mortem transported individual shells (Urlichs & Kurzweil 1997) or active immigration attempts by Alpine ammonoids?
According to Brack & Rieber (1993), Flexoptychites angusto-umbilicatus, as described by Urlichs & Kurzweil (1997), is a common species that continues through to the Anisian/Ladinian transition in the secadensis zone without significant morphological change. According to Urlichs (1978), Protrachyceras recubariense would be interpreted as a problematic species of the curionii zone, as it does not morphologically correspond to any known form. In contrast, Brack & Rieber (1993) and Brack et al. (1995, 1996, 1999) place the morph as Nevadites sp. in the secadensis zone. Mietto et al. (2018), in turn, identify the morph as C. pemphix (Merian) from the interval between the chiesense subzone and the basal part of the curionii subzone.
In the Southern Alpine Buchenstein Formation, the – GSSP – Anisian/Ladinian boundary is located at the transition from the secadensis to the curionii zone (Storck et al. 2018). In the view of specialists, the Flexoptychites and Protrachyceras of the SW German spinosus zone originate from this stratigraphically significant boundary region. They therefore hold a special value status for the chronostratigraphic correlation between the Germanic and Alpine Middle Triassic. Two variants regarding the immigration timing of the Alpine morphs can be discussed.
Variant 1: With a further temporal immigration window, the Anisian/Ladinian boundary in the Upper Muschelkalk would thus be shifted to the middle spinosus zone, and the fossil occurrence would chronostratigraphically and biostratigraphically correspond to the Buchenstein Formation. The situation could be causally attributed to post-mortem transported individual shells or a brief, unsuccessful immigration attempt by Alpine ammonites.
Variant 2: If the Southern Alpine ammonites immigrated concurrently with the brachiopod Punctospirella fragilis, post-mortem transport of individual shells would be ruled out. The timing of the fourth immigration phase would chronostratigraphically and biostratigraphically correspond to a successful immigration attempt of entire populations at the Anisian/Ladinian transition. Following territorial and reproductive isolation, the immigrated ammonites developed into distinct cladogenetic species up into the spinosus zone. The conservative Flexoptychites remained without, and Protrachyceras with unknown morphological injury on the venter, based on which Spath (1934) removed the specimen from the genus Ceratites de Haan 1825 and unjustifiably placed it in its own genus Alloceratites (an injury does not justify a taxonomic change).
Abb. 6: Cerattes schmiat und Neoclypues . peregrinus
a) Ceratites schmidi, Original Philppi 1901. Abb. 5 und Sa. aa) Ceratites schmidi, Orginal Philipp 1901. Abb. Sb und Sc. b) Ceratites schmidi Gipsabgusse vom zweiten rund 196l (palaont. Sammlung Halle Nr. H 61). Museum TU Freiberg Nr. 123/1, C) Ceratites schmidi, Original vom dritter Fund 1970, paläont. Sammlung TU Freiberg Nr. 144/1, d) Neoclypites ? peregrinus, Original, paläont. Sammlung TU Freiberg Nr. 144/2
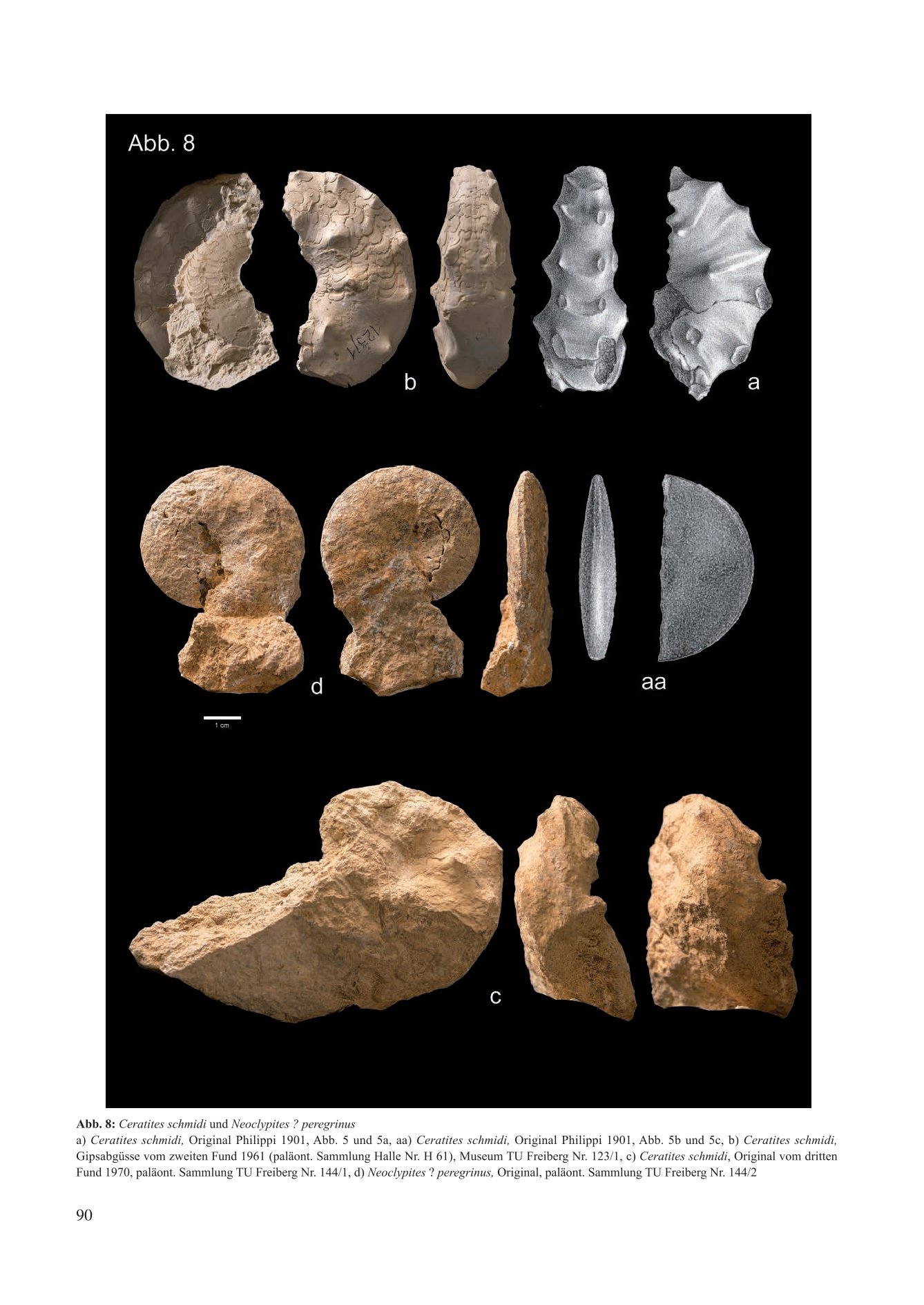
With the discovery of a second ceratite from the Thuringian Grenzdolomit in the Reisdorf brickyard pit near Apolda (1961), a taxonomic problem arose, as the new specimen possesses a sculpture-free, smooth venter (Fig. 8 c/d). To avoid creating a new species and a new subgenus for the Reisdorf find, Müller (1969) subsequently reassigned Ceratites schmidti to the genus Ceratites de Haan and relegated the taxon Alloceratites Spath, 1934 to the rank of a subgenus.
With further new finds from the Grenzdolomit of the Reisdorf brickyard pit (Fig. 8 e/f), the taxonomic assignment changed once again (Müller 1973). The first specimen represents the "normal form" of a Ceratites schmidti. The second has a flat, disc-shaped shell with a furrow running partially medially along the venter. It is described as the new species Neoclypites ? peregrinus n. sp. Kozur (1971) assumes that Israelites ramonensis from the Ladinian of Israel shares certain characteristics with both morphs and suspects, due to chronostratigraphic factors, closer phylogenetic relationships between Alloceratites and Hungarites than with Ceratites. Thus, Müller (1973) finally detached Ceratites schmidti from the genus Ceratites de Haan and assigned the ceratites from the Thuringian Grenzdolomit solely to Alloceratites Spath, 1934.
With the cladogenetic splitting of a population of the genus Ceratites de Haan into the sister species Ceratites tornquisti and Ceratites nodosus, the genealogical origin is clarified (Fig. 2c). The endemic phylogenesis of the evolutionary species Ceratites nodosus ends in the Upper Muschelkalk at the base of the Erfurt Formation with the extinction of the species. Parallel to this, the heterogeneous evolution of the sister species proceeds spatially and reproductively separated within the Palaeotethys. Regardless of further speciation events during phylogenesis, all descendants of the species Ceratites tornquisti remain part of the genus Ceratites de Haan. Therefore, the immigration in the Longobardian occurred from an unknown refugium of the Palaeotethys as the species Ceratites schmidti (Fig. 3c).
Only the interpretation of the median furrow of "Neoclypites ? peregrinus" remains uncertain. The shell was originally embedded ventrally in a granular dolomitic block. Only if the furrow was created mechanically through breakage would it represent a juvenile individual of the genus Ceratites de Haan and a descendant of the original sister species Ceratites tornquisti. Otherwise, the genealogy of the species Neoclypites ? peregrinus remains unknown.
Another problem is posed by the migration of Germanonautilus jugatonodosus into the Grenzdolomit. According to Merki (1961), the Upper Muschelkalk in the eastern Swiss Jura ended with the equivalent of the Grundgips. Accordingly, the southern marginal area of the Muschelkalk sea in northern Switzerland would have become a retreat for the marine Muschelkalk fauna, including Germanonautilus bidorsatus (Rein 2014). Since Pietsch et al. (2016) corrected the stratigraphic range of the Lower Keuper, the immigration of Germanonautilus jugatonodosus must be revised without reference to the phylogenesis of Germanonautilus bidorsatus (Urlichs 2000, 2015, Rein 2015).
8. Conclusion
As a newly emerged ecosystem, the Upper Muschelkalk represents a tectonically dominated epicontinental sea characterized by its constantly changing marine connections to various faunal provinces of the Palaeotethys and Neotethys. Immigrated macrofossils enable biostratigraphic and chronostratigraphic correlation as well as the reconstruction of hypothetical migration routes into the inland sea, chronologically marked by Germanonautilus bidorsatus and Sephardonautilus tridorsatus and their respective hypothetical gateways.
The first to migrate, during the lower trinodosus zone, was Germanonautilus salinarius from the Austroalpine-Dinaric faunal province of the Palaeotethys via a hypothetical Helvetic-Alemannic gateway into the basin interior. The species Germanonautilus bidorsatus, which emerged cladogenetically from the ancestral species, succeeded in physiologically adapting to all changing ecosystems from the Diemel Formation to the Erfurt Formation.
From the Austroalpine faunal province, Sturia sansovinii migrated along the same route into the basin interior during the middle trinodosus zone, cladogenetically forming the new species Sturia brandti n. sp.
The most significant step in the colonization of the Upper Muschelkalk ecosystem occurred in the lower Illyrian from the Sephardic faunal province. Through a uniquely and briefly tectonically created Burgundian-Rhenish gateway, alien cephalopods from the Neotethys migrated into the inland sea. With Sephardonautilus tridorsatus nov. comb. as the type species of the new genus Sephardonautilus and the ancestral species Sephardonautilus salinarius, the phylogenetic relationship between the individuals of the Muschelkalk sea and the Sephardic faunal province is documented.
The same applies to the evolutionary species Ceratites nodosus, which emerged as a sister species through allopatric speciation from an unknown species of the genus Ceratites de Haan of the Sephardic faunal province. The ancestral species Sephardonautilus salinarius from the trinodosus zone of the upper Gevanim Formation of Makhtesh Ramon confirms the chronological correlation of the migration route of the cladogenetically emerged Sephardonautilus tridorsatus in the Upper Muschelkalk. The morphocline of the unprecedentedly eurypotent evolutionary species Ceratites nodosus allows for a seamless biostratigraphic subdivision of the Upper Muschelkalk.
With the chronological correlation of the ancestral species of Germanonautilus bidorsatus and Sephardonautilus tridorsatus to the Austroalpine and Sephardic faunal provinces, the trinodosus zone becomes the stratigraphic basis of the Upper Muschelkalk.
All immigrated Alpine ammonites prove to be only partially physiologically adaptable compared to the eurypotent species Ceratites nodosus. The immigrants Pinacoceras and Gymnites from the Lower Ladinian Schreyeralm Formation reach the basin center. However, the physiological integration capacity of the cladogenetically emerged species Parapinacoceras thiemei and Gymnites bruneli remains limited.
The adaptive behavior of the Southern Alpine morphs Flexoptychites angusto-umbilicatus and Protrachyceras recubariense appears surprising. It is difficult for several reasons to view them as passively transported, isolated shell remains in the spinosus zone of SW Germany. A second active variant is favored. If immigration occurred concurrently with the brachiopod Punctospirella fragilis, post-mortem transport of individual shells is ruled out. The timing of the fourth immigration phase would chronostratigraphically and biostratigraphically correspond to a successful immigration attempt of entire populations at the Anisian/Ladinian transition. The immigrated Flexoptychites and Protrachyceras survived in the SW German region with physiological adaptation into the spinosus zone. During this time, they cladogenetically acquired a new species status.
For the definition of the Anisian/Ladinian boundary, the Spiriferina bed would mark the transition from the compressus zone to the evolutus zone. This correlates with the transition from the secadensis zone to the curionii zone in the Southern Alpine faunal province.
The evolution of the unknown species of the genus Ceratites de Haan from the Sephardic faunal province ends with allopatric splitting into the two sister species Ceratites nodosus and Ceratites tornquisti. The phylogenesis of the species Ceratites tornquisti began in the lower Illyrian in the Palaeotethys and ended in the Longobardian within the Grenzdolomit in Thuringia as Ceratites schmidti. Due to its fastigate sculpture, C. schmidti was removed from the genus Ceratites de Haan by Spath (1934) and unjustifiably placed in its own genus Alloceratites. A traumatically induced morphological change in an individual does not alter a generic status. With the clarification of the genealogical origin of Ceratites tornquisti, the sister species of Ceratites nodosus, all descendants of the species remain part of the genus Ceratites de Haan. The clarification of the previously unknown phylogenesis of the species Ceratites tornquisti in the habitats of Dobruja, Sardinia, Vicentin, Provence, and Thuringia remains reserved for detailed analysis.
Acknowledgements
I would like to thank Mr. M. Hartmann (Natural History Museum Erfurt) for his multifaceted support of my work, Prof. Dr. F. Hirsch (Naruto University) for his expert advice, Mr. S. Brandt (Kornhochheim) for the photography, Dr. J. A. Perez Valera (University of Madrid) for his assistance in procuring important literature, and Prof. Dr. J. W. Schneider (TU Freiberg) for the loan of the original Ceratite specimens from the Grenzdolomit.
Literature
Adams, A. & L. W. Diamond (2019): Facies and depositional environments of the Upper Muschelkalk (Schinznach Formation, Middle Triassic) in northern Switzerland. – Swiss Journal of Geosciences, 25 S., 14 Abb.
Anastasiu, V. (1898): Contribution ix l’etude geologique de la Dobrogea. Terrains secondaires. – These pres. a la faculte des Sciences de Paris, p. 46.
Bachmann, G. H., M. Geluk, J. Warrington, A. Becker, G. Beutler, H. Hagdorn, M. Hounslow, E. Nitsch, H.-G. Röhl, Th. Simon & A. Szulc (2010): with contributions of Dusar, M., Nielsen, L. H., Barnasch, J., Franz, M.: Triassic. – In: Doornenbal, H. & A. Stevenson (eds.): Petroleum Geological Atlas of the Southern Permian Basin Area, 149–173; Houten/The Netherlands (EAGE).
Bittner, A. (1890): Brachiopoden der alpinen Trias. – Abhandlungen der kaiserlich-königlichen geologischen Reichsanstalt XIV, 325 S., 41 Taf.
Böttcher, R. J. (1938): Versteinerungen des Oberen Muschelkalks bei Ohrdruf als aufschlussreiche Dokumente für die Geschichte des deutschen Muschelkalkmeeres. – Beiträge zur Geologie von Thüringen 5: 99–105.
Brack, P. & H. Rieber (1993): Towards a better definition of the Anisian/Ladinian boundary: New biostratigraphic data and correlations of boundary sections from the Southern Alps. – Eclogae geologicae Helvetiae 86/2: 415–527, 17 Abb., 14 Taf.
Brack, P., H. Rieber & M. Urlichs (1999): Pelagic successions in the Southern Alps and their correlation with the Germanic Middle Triassic. – Zentralblatt Geologie und Paläontologie Teil I, Heft 7–8, 853–976, 8 Fig., Stuttgart.
Brack, P., H. Rieber & R. Mundil (1995): The Anisian/Ladinian Boundary interval at Bagolino (Southern Alps, Italy): In. Summary and news results on ammonoid horizons and radiometric age dating. – Albertiana 15: 45–56, 3 Fig.
Brack, P., H. Rieber & M. Urlichs (1996): The new „high resolution Middle Triassic ammonoid standard scale“ proposed by Triassic researchers from Padova – a discussion of the Anisian/Ladinian boundary interval. – Albertiana 17: 42–50.
Brandt, S. & S. Rein (2019): Die Chronospezies Ceratites flexuosus von Geilsdorf – Ein evolutionsbiologisches Modellfall der evolutionären Art Ceratites nodosus. – VERNATE 38: 53–76.
Broili, F. (1927): Eine Muschelkalkfauna aus der Nähe von Saalfelden. – Sitzungsberichte der mathematisch-naturwissenschaftlichen Abteilung der BAK, Heft III: 229–242.
Diener, C. (1900): Die triadische Cephalopodenfauna der Schichlinghöhe bei Hallstatt. – in: Beiträge zur Paläontologie und Geologie Österreich-Ungarns und des Orients; Ed. Dr. G. v. Arthaber; Band XIII, Wien und Leipzig 1901; Seite 3–42 und 3 Tafeln.
Diener, C. (1916): Einige Bemerkungen zur Nomenklatur der Triascephalopoden. – Centralblatt für Mineralogie, Geologie und Paläontologie 1916: 97–105.
De Haan, G. (1825): Monographiae Ammoniteorum et Goniatiteorum Specimen. – II + 168 S.; Leyden (Hazenberg).
Hagdorn, H. & T. Simon (1993): Ökostragraphische Leitbänke im Oberen Muschelkalk. – In: Hagdorn, H. & A. Seilacher (eds.): Muschelkalk. Schöntaler Symposium 1991, Stuttgart, Korb (Goldschneck); 193–208, 15 Abb.
Hauer, F. v. (1887): Die Cephalopoden des Bosnischen Muschelkalkes von Han Bulog bei Sarajevo. – Denkschrift der mathematisch-naturwissenschaftlichen Classe der kaiserlichen Akademie der Wissenschaften, 8 Taf.
Hauer, F. v. (1892): Cephalopoden aus der Trias von Bosnien I. – Denkschrift der mathematisch-naturwissenschaftlichen Classe der kaiserlichen Akademie der Wissenschaften, 15 Taf.
Hauer, F. v. (1896): Cephalopoden aus der Trias von Bosnien II. – Denkschrift der mathematisch-naturwissenschaftlichen Classe der kaiserlichen Akademie der Wissenschaften, 13 Taf.
Hohenstein, V. (1913): Beiträge zur Kenntnis des Mittleren Muschelkalks und des unteren Trochitenkalks am östlichen Schwarzwaldrand. – Geologisch-paläontologische Abhandlungen (16), N.F.12: 175–272, 12 Abb., Taf. 12–19.
Knaust, D. (1991): Ein bemerkenswerter Nautiliden-Kiefer (Rhyncholites hirundo Faure-Biguet, Cephalopoda) aus dem Mittleren Muschelkalk (Oberanis) der Germanischen Trias. – Veröffentlichungen des Naturkundemuseums Erfurt 10: 58–65, 4 Abb., 1 Taf.
Kozur, H. (1971): Mikropaläontologie, Biostratigraphie und Biofazies der germanischen Mitteltrias. – Freiberg Bergakademie, Dissertation, Freiberg.
Kozur, H. (1974): Biostratigraphie der germanischen Mitteltrias, Teil I und II mit Anlagen. – Freiberg Forschungshefte C, 280, I: 1–56, II: 1–71.
Kummel, B. (1960): Middle Triassic Nautiloids from Sinai, Egypt and Israel. – Bulletin of the Museum of Comparative Zoology at Harvard College 123 (7): 291–304, 2 Pls.
Merki, P. J. (1961): Der Obere Muschelkalk im östlichen Schweizer Jura. – Birkhäuser AG, 263 S., Basel.
Mietto, P., S. Manfrin & M. Rigo (2018): Middle Triassic ammonoid fauna from the Recoaro and Tretto areas (NE Italy) and its stratigraphic and paleobiogeographic evidence. – Bollettino della Società Paleontologica Italiana 53 (3), 217–250, 8 Fig.
Mojsisovics, E. v. (1873–1902): Das Gebirge um Hallstatt; Abt. I, Die Cephalopoden der Hallstätter Kalke. – Abhandlungen der kaiserlich-königlichen Geologischen Reichsanstalt 6: 1–356, pl. 1–94.
Mojsisovics, E. v. (1882): Cephalopoden der mediterranen Triasprovinz. – Abhandlungen der kaiserlich-königlichen Geologischen Reichsanstalt 10: 1–322, 1–94.
Mojsisovics, E. v. (1902): Das Gebirge um Hallstatt. – Abhandlungen der kaiserlich-königlichen Geologischen Reichsanstalt, 3. Lief. (Supplement): 175–356, Taf. 1–23.
Müller, A. H. (1969): Ein Ceratit (Ceratites cf. schmidti, Ammonoidea) aus dem Unterkeuper (Grenzdolomit) des Germanischen Triasbeckens. – Monatsberichte der Deutschen Akademie der Wissenschaften zu Berlin 11, 2: 122–132, 5 Abb., 2 Taf.
Müller, A. H. (1970): Neue Funde seltener Ceratiten aus dem germanischen Muschelkalk und Keuper. – Monatsberichte der Deutschen Akademie der Wissenschaften zu Berlin 12, 8: 632–642, 16 Abb., 1 Taf.
Müller, A. H. (1973): Über Ammonoidea (Cephalopoda) aus dem Grenzdolomit des germanischen Unterkeupers. – Zeitschrift für geologische Wissenschaft 1, 8: 935–945, 9 Abb., 1 Taf.
Pietsch, J. S., Wetzel, A. Jordan, P. (2016): A new lithostratigraphic scheme for the Schinznach Formation (upper part of the Muschelkalk Group of northern Switzerland). – Swiss Journal of Geosciences: 285–307, 15 Fig.
Reif, W. E. (2009): Artabgrenzung und das Konzept der evolutiven Art in der Paläontologie. – Journal of Zoological Systematics and Evolutionary Research 22 (3): 263–286, 2 Abb.
Rein, S. (2007): Die Biologie der Ceratiten der flexuosus-, sequens und pulcher und semipartitus/meissnerianus-Zone. – Entstehung und Aussterben der Biospezies Ceratites nodosus. – Veröffentlichungen des Naturkundemuseums Erfurt 26: 39–67, 32 Abb., 6 Taf., 3 Prof.
Rein, S. (2008): Ceratites avitus auf Mallorca? – Die Muschelkalkammonoideen des Museu Balear de Sciencies Naturals Sóller. – VERNATE 27: 5–19, 7 Abb.
Rein, S. (2009): Stratigraphie und Fossilführung des Oberen Muschelkalkes bei Eisenach und Jena. – VERNATE 28: 31–49, 10 Abb., 5 Taf.
Rein, S. (2014): Germanonautilus im Unteren Keuper (Trias, Erfurt-Formation) – von Trematodiscus jugatonodosus bis Germanonautilus bidorsatus. – Semana 29: 37–44, 11 Abb.
Rein, S. (2015): Erwiderung auf Urlichs vorstehende „Entgegnung zu Rein (2014)“: Germanonautilus im Unteren Keuper (Trias, Erfurt-Formation) – von Trematodiscus jugatonodosus bis Germanonautilus bidorsatus. – Semana 30: 57–58, 1 Abb.
Rein, S. (2016): Germanonautilus bidorsatus und ?Germanonautilus tridorsatus (Nautiloidea) aus dem Oberen Muschelkalk (Mitteltrias, Anis–Ladin). Teil 1: Speziation, Ontogenese und Phylogenese. – VERNATE 35: 31–65, 13 Abb., 11 Taf.
Rein, S. (2017): Speziation, Phylogenie und Ontogenie der Biospezies Ceratites nodosus und das Chronospezies Konzept. – VERNATE 36: 47–64, 18 Abb.
Rein, S. & R. Werneburg (2010): Parapianoceras und Gymnites (Ammonoidea) aus der enodis/posseckeri-Zone im Oberen Muschelkalk (Mittel-Trias, Ladin) Thüringens. – Semana 25: 87–100, 9 Abb., 1 Taf.
Seidel, G. (2013): Stratigraphie, Fazies und geologische Stellung des Zechsteins und der Trias Thüringens. – Beiträge zur Geologie von Thüringen N.F. 20: 21–78, 27 Abb., 12 Tab.
Schlotheim (1820): Petrefaktenkunde. – Gotha.
Schmidt, M. (1928): Die Lebewelt unserer Trias. – Hohenloh’sche Buchhandlung F. Rau; 461 S., 2300 Abb., Öhringen.
Schröder, W. (1977): Ein besonderer Fund … – Der Aufschluss 28, 110, 1 Abb.
Stampfli, G. M. & G. D. Borel (2002): A plate tectonic model for the Paleozoic and Mesozoic constrained by dynamic plate boundaries and restored synthetic oceanic isochrones. – Earth and Planetary Science Letters 196: 17–33.
Schnetzer, R. (1934): Die Muschelkalkfauna des Ofenbachgrabens bei Saalfelden. – Palaeontographica 81: 1–160, 15 Textabb., Taf. 1–6.
Spath, L. F. (1934): The Ammonoidea of the Trias (II). – Catalogue of Fossils, Ceph. Brit. Mus. IV. 521 S., 18 Taf.
Spath, L. F. (1946): The Middle Triassic Cephalopoda from Sinai. – Institute Egypt Bulletin 27: 425–426, pl. III.
Storck, J.-C., P. Brack, J.-F. Wotzlaw & P. Ulmer (2018): Timing and evolution of Middle Triassic magmatism in the Southern Alps (northern Italy). – Journal of the Geological Society; 16 S., 12 Fig., 1 Tab.
Szulc, J. (2000): Middle Triassic evolution of the northern Peri-Tethys area as influenced by early opening of the Tethys Ocean. – Annales Societatis Geologorum Poloniae 70: 1–48.
Tollmann, A. (1976): Analysen des klassischen nordalpinen Mesozoikums. – Franz Deuticke Wien, 581 S.
Tornquist, A. (1898): Neue Beiträge zur Geologie und Paläontologie der Umgebung von Recoaro und Schio (im Vicentin). 2. Beitrag: Die Subnodusus-Schichten. – Zeitschrift der deutschen Geologischen Gesellschaft 50: 635–694, Taf. XX–XXIII.
Tornquist, A. (1901): Das Vorkommen von nodosen Ceratiten auf Sardinien und über die Beziehungen der mediterranen zu den deutschen Nodosen. – Centralblatt Mineralogie etc.: 385–396